|
| Appunti universita |
|
|
| Appunti universita |
|
| Visite: 2530 | Gradito: |
Leggi anche appunti:Ipertensione cronica in gravidanzaIpertensione cronica in Gravidanza Definizione di Ipertensione Cronica (HCR) Ipertensione Medioevo e rinascimentoMEDIOEVO E RINASCIMENTO Il medioevo Il medioevo è un periodo storico che Disturbi alimentari e assistenza al malatoDisturbi alimentari e assistenza al malato I disturbi alimentari Per |
 |
 |
Recettore adrenergico
Struttura recettoriale
Il gene umano che codifica per il recettore β -adrenergico (β -AR) è situato sul braccio lungo del cromosoma 5 ed è costituito da 1.200 paia basi (Kobilka et al.,
Il recettore è una proteina di 46,5 Kda costituita da 413 residui amminoacidici (Kobilka et al., 1987), distribuiti in 7 domini transmembranari tra i quali se ne riconoscono 3 extracitoplasmatici, con la porzione ammino-terminale, e 3 intracitoplasmatici, con la porzione carbossi-terminale. Il recettore viene attivato mediante N-glicosilazione sugli amminoacidi 6, 15 e 187; un processo importante non solo per l'inserimento all'interno della membrana cellulare, ma anche per il legame del recettore con l'agonista (Johnson, 2006). La presenza dell'amminoacido cisteina in posizione 341 è fondamentale per l'ancoraggio della catena carbossi-terminale con la membrana citoplasmatica (O'Dowd et al., 1989).
2 Attivazione del recettore e trasduzione del segnale
L'attivazione
del recettore β -AR comporta l'incremento intracellulare dei livelli di cAMP (Robison et al., 1967). Questo è il risultato della stimolazione
dell'adenilato ciclasi, che catalizza la conversione dell'adenosina trifosfato (ATP) in cAMP. L'attivazione della proteina trimerica Gs si ottiene tramite il legame tra
recettore β e adenilato ciclasi (Figura 6).
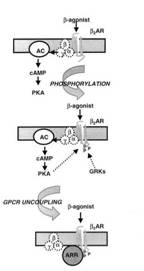
Fig Attivazione recettore AR
La proteina G è costituita da una subunità α (che stimola l'adenilato ciclasi) e le subunità βγ (che trasducono il segnale). I livelli di cAMP, sono regolati dalle fosfodiesterici, che degradano il cAMP in 5'-AMP. Il meccanismo tramite il quale cAMP determina il rilassamento della muscolatura liscia respiratoria non è del tutto chiaro, ma sembra ciò si realizzi mediante l'attivazione della proteina chinasi A (PKA), che a sua volta fosforila proteine-chiave coinvolte nel processo di contrazione muscolare (Johnson et al., 1995). Il cAMP inoltre, risulta essere un inibitore del rilascio del calcio (Ca dai depositi intracellulari, inibendone anche l'ingresso nella cellula e portando ad un rilassamento della muscolatura liscia. Comunque è stato visto recentemente che alcune risposte indotte dai β agonisti sono mediate da meccanismi cAMP-indipendenti, che coinvolgono direttamente la proteina Gs e i canali al potassio (K+) presenti nelle cellule muscolari lisce dell'albero respiratorio (O'Dowb et al., 1989).
I recettori β2 possono essere accoppiati anche a proteine Gi (Daaka et al., 1997). Il risultato di questa associazione è la stimolazione di un segnale intracellulare che porta all'attivazione della p38MAPK, una proteina chinasi coinvolta nella trasmissione del segnale mitogenico. Per l'attivazione di questa via è necessaria la fosforilazione PKA-mediata del recettore β , l'assemblaggio di proteine intracellulari come Raf, Csrc, RAS con la subunità βγ della proteina G, e l'attivazione di MAPK (Daaka et al., 1997).
Da studi recenti è emerso, inoltre, che l'attivazione di questa via porta alla fosforilazione, da parte della MAPK, del recettore glucocorticoide (GR) rendendolo più sensibile all'azione degli steroidi (Johnson, 2002). L'attivazione del recettore β -AR porta anche alla traslocazione del recettore GR dal citoplasma al nucleo, uno dei passaggi fondamentali nel meccanismo d'azione di questi farmaci (Roth et al., 2002).
3 Desensibilizzazione recettoriale
Associato con l'attivazione del recettore β -adrenergico vi è il processo autoregolatorio di desensibilizzazione recettoriale. Questo processo avviene fisiologicamente per preservare il recettore da una sovrastimolazione indotta da agonisti endogeni ed esogeni. La desensibilizzazione avviene in seguito al legame tra recettore e agonista ed i meccanismi attraverso i quali può avvenire sono di 3 tipi (Figura 7): (1) disaccoppiamento tra recettore e adenilato ciclasi, (2) internalizzazione del recettore non accoppiato e (3) down-regulation.
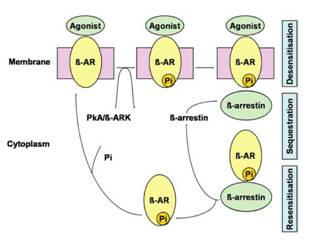
Fig . Desensibilizzazione del recettore AR.
Il legame tra recettore e agonista innesca quasi immediatamente la fosforilazione di specifici residui amminoacidici di serina e treonina nel terzo loop intracitoplasmatico e nella terminazione carbossi-terminale del recettore. Il β -AR è fosforilato dalla PKA o da chinasi specifiche delle proteine G (GRK) (Bouvier at al., 1988, 1989; Hausdorff et al., 1989).
Il legame dell'agonista con il recettore porta ad una immediata traslocazione del GRKs dal citoplasma alla membrana e alla successive fosforilazione del recettore. Una volta ultimata la fosforilazione, la β-arrestina si lega al recettore, non permettendo più l'accoppiamento del recettore con la proteina Gs, limitando così la funzionalità recettoriale stessa. L'azione della β-arrestina si esplica anche mediante il legame con altre proteine, come la fosfodiesterasi IV (Johnson, 2006).
L'attivazione di altri recettori, che utilizzano cAMP come secondo messaggero, possono indurre desensibilizzazione eterologa del recettore β -AR, a causa della formazione di cAMP e della successiva attivazione di PKA. La desensibilizzazione omologa, invece, è la risultante dell'attivazione combinata di PKA e GRK.
L'arrestina, non svolge solo un importante ruolo nel disaccoppiamento recettoriale, ma è anche un mediatore cruciale coinvolto nell'internalizzazione del recettore, infatti vi è una forte connessione tra la fosforilazione del recettore β - AR e il legame della β-arrestina con il processo di endocitosi di β -AR. Una delle funzioni della β-arrestina è quella di collegare il recettore β -AR con il sistema di endocitosi (Goodman et al., 1997; Laporte et al., 1999, 2000). In seguito all'internalizzazione, il recettore β -AR può andare incontro a defosforilazione da parte degli enzimi endosomiali ed essere riciclatato sulla membrana cellulare, oppure può essere degradato dai lisosomi (Campbell et al., 1991; Barak et al.,
1994). L'internalizzazione e la degradazione lisosomiale del recettore β -AR comportano la necessità della neosintesi del recettore, affinché venga nuovamente espresso.
3. La down-regulation del recettore β -AR è un processo che porta alla desensibilizzazione, come risultato di eventi trascrizionali. Lo studio condotto da Hadcock e Malbon (1988) ha dimostrato che in seguito all'esposizione all'agonista forskolina, alla concentrazione 10 µM per 18 ore, si ha una riduzione del 50% dei livelli di espressione del recettore.
Oltre alla capacità dei β-agonisti di indurre desensibilizzazione omologa, altri stimoli sono capaci di ridurre la risposta β-adrenergica, un processo definito desensibilizzazione eterologa. Ad esempio, gli agonisti muscarinici hanno la capacità di attivare la subunità inibitrice Giα delle proteine G portando ad un'inibizione dell'adenilato ciclasi e attenuando la risposta del β-agonista attraverso l'attivazione di PKC (Grandorby et al., 1994). Inoltre, uno studio recentemente pubblicato da Ahiua e collaboratori (2008), ha dimostrato che la desensibilizzazione eterologa dei recettori β -AR, risultante da una esposizione prolungata alla PGE2 o alla forskolina, è mediata da un aumento dell' attività
della PDE4. Numerose citochine, quali IL-1β, TNFα, IL-5 e TGFβ riducono la capacità delle cellule muscolari bronchiali lisce di generare cAMP. Le citochine, non alterano l'espressione della PKA e non hanno effetto sulla risposta cellulare alla forskolina, un agente che attiva direttamente l'adenilato ciclasi (Shore et al.,
1997; Laporte et al., 1998; Pang et al., 1998; Pascual et al., 2001). Questi risultati suggeriscono che il meccanismo di desensibilizzazione, in questo caso, si realizzi a livello dell'accoppiamento tra recettore β-adrenergico e proteina Gs (Shore et al., 1997).
|
| Appunti su: recettore appunti universitari, reccertori CEB22 aunento cAMP, |
|
| Appunti Bambini |  |
| Tesine Nutrizione |  |
| Lezioni Bellezza |  |