|
| Appunti superiori |
|
|
| Appunti superiori |
|
| Visite: 3685 | Gradito: |
Leggi anche appunti:I fioriI fiori Obiettivo: osservazione dell'apparato riproduttore dei fiori Strumenti/attrezzatura: Ricerca di Biologia - Condritti & OsteittiRicerca di Biologia Condritti & Osteitti Introduzione I pesci, sono animali Le caratteristiche degli esseri viventiLe caratteristiche degli esseri viventi 1. Hanno strutture complesse e organizzate, |
 |
 |
Tronco encefalico
Descrizione
Il tronco encefalico è la parte del SNC che connette il midollo spinale al cervello. In realtà essendo il SNC un continuum non ci sono limiti o zone di confine nette che indicano dove finisce il midollo e dove inizia il bulbo. A dimostrazione di ciò alcuni nervi encefalici che hanno i loro nuclei per lo più nel tronco encefalico hanno anche alcuni nuclei, o la continuazione di alcuni nuclei, nella parte più rostrale del midollo spinale.
Si considera tronco encefalico la parte del SNC che al livello del grande forame occipitale continua dal midollo fino al diencefalo. Posteriormente il tronco encefalico è notevolmente abbarbicato al cervelletto (anche perchè condividono un'unica origine dal punto di vista embriologico), adagiato in parte sulla parte basilare dell'osso occipitale (bulbo) e in parte sulla faccia posteriore del corpo dello sfenoide e sul dorso della sella (ponte) nella fossa cranica posteriore.
Il cervelletto è connesso al tronco encefalico tramite tre fasci di fibre che formano i peduncoli cerebellari, i peduncoli cerebellari inferiori, medi e superiori.
Lungo tutto il tronco encefalico emergono 10 dei 12 nervi cranici.
Il tronco dell'encefalo è diviso in tre parti in senso rostro-caudale:
-il mesencefalo
-il ponte
-il bulbo o midollo allungato.
Queste parti embriologicamente, come tutto il sistema nervoso, derivano dal tubo neurale. La parte più superiore, derivante dalla prima piega del tubo neurale, è chiamata mesencefalo. Caudalmente al mesencefalo il tubo neurale, che sopra si chiude a formare la lamina del tetto, presenta un difetto di chiusura per cui le due superfici dorsali del tubo non si chiudono e lasciano una struttura aperta a forma romboidale, il romboencefalo. Il romboencefalo dà vita cranialmente al ponte, detto metencefalo a livello organogenetico, e caudalmente al bulbo, detto mielencefalo.
A sviluppo completato, in direzione ventro-dorsale, il mesencefalo presenta una superficie ventrale e un tetto, il ponte e parte del bulbo invece mancano del tetto. Nel mesencefalo tra la superficie ventrale e il tetto scorre un canale che mette in comunicazione il 3° ventricolo con il 4° ventricolo, l'acquedotto mesencefalico (o di Silvio). Tale acquedotto estua nel 4° ventricolo che è pertanto situato a livello del ponte e, in parte, del bulbo.
Bulbo
Il bulbo visto dal davanti presenta in alto, al confine con il ponte, un solco netto che lo divide da esso, il solco bulbo pontino (il bulbo è diviso dal ponte perché il ponte, a differenza delle altre componenti del tronco encefalico, ha solo uno sviluppo orizzontale).
Procedendo in direzione caudale, sul davanti compaiono dei rilievi cilindrici che, per il loro aspetto trapezoidale in sezione, sono chiamati piramidi; sono divise da un solco mediano, che poi nel midollo continua come solco mediano anteriore. Tale solco, nella parte più inferiore delle piramidi è completamente obliterato dalla decussazione delle piramidi, ovvero dall'imponente passaggio di fibre (circa l'80%) che avviene da una piramide all'altra, le quali fibre nel midollo costituiscono, nel cordone dorso-laterale, la via cortico-spinale o piramidale. Poiché non tutte le fibre decussano, al di sotto della decussazione delle piramidi alcune fibre (circa il 20%) formano nel cordone anteriore del midollo spinale la via piramidale ventrale.
Lateralmente e posteriormente alle piramidi, nella parte più superiore del bulbo, si rileva la presenza, uno per lato, di altri due rilievi che per la forma ovalare prendono il nome di olive inferiori (o bulbari). Posteriormente alle olive altri due rilievi formano i peduncoli cerebellari inferiori.
Le olive sono un importante punto di riferimento perché determinano due solchi, uno anteriormente, il solco preolivare, e uno posteriormente, il solco retroolivare. Da questi solchi si nota l'emergenza di alcuni nervi cranici.
Lateralmente al bulbo si estendono i recessi del quarto ventricolo (delle saccocce derivate da un'estensione del velo midollare). A livello dei recessi c'è una formazione spugnosa (lungo tutto il sistema cefalo rachidiano c'è la presenza di queste formazioni), formata da plessi vascolari, in cui si forma il liquido cefalo rachidiano (che si forma anche già nell'embrione, nel canale neurale). Anteriormente i recessi presentano dei fori (di Luschka) che permettono al liquor di accedere allo spazio subaracnoideo.
Dorsalmente il bulbo è coperto dal cervelletto. Asportato il cervelletto il bulbo non è delimitato dal ponte tramite un solco, come lo è anteriormente. Con l'asportazione del cervelletto vengono sezionati i tre peduncoli cerebellari e pertanto posteriormente come limite superiore del bulbo si fa riferimento ad un'ipotetica linea passante dal limite inferiore dei peduncoli cerebellari medi.
La parte dorsale del bulbo presenta una componente rostrale e una componente caudale. La componente rostrale del bulbo partecipa a formare il pavimento del quarto ventricolo. É aperta a forma di V ed è formata lateralmente dai peduncoli cerebellari inferiori. L'apice della V forma l'obex in corrispondenza del quale si trova l'area postrema (zona dell'innesco del vomito).
Al di sotto della V si estende la parte caudale del bulbo che presenta medialmente i rilievi bilaterali dei nuclei gracile e lateralmente i nuclei cuneato e cuneato laterale in cui terminano i rispettivi fascicoli.
Ponte
Il ponte si estende rostralmente e anteriormente al di sotto dei peduncoli cerebrali mentre rostralmente e posteriormente al di sotto del tetto del mesencefalo. Tra mesencefalo e ponte si trova la parte più stretta del nevrasse detta istmo ponto-mesencefalico. Anteriormente il ponte presenta la caratteristica forma da cui prende il nome. Questa parte è detta parte basilare del ponte e presenta sulla linea mediana un solco in cui decorre l'arteria basilare. La parte basilare è formata, tra le altre, da fibre che si portano al cervelletto per cui dorsalmente forma i peduncoli cerebellari medi.
La parte dorsale del ponte è formata dal tegmento pontino e presenta lateralmente parte dei peduncoli cerebellari superiori. Partecipa, insieme alla parte rostrale del bulbo, alla composizione del pavimento del 4° ventricolo.
Quarto ventricolo
Del quarto ventricolo si considerano un pavimento, due pareti laterali e un tetto. Le pareti laterali sono date dai peduncoli cerebellari inferiori. Il tetto è formato nella parte rostrale dai peduncoli cerebellari superiori mentre nella sua parte caudale, non avendo ponte e il bulbo nessuna chiusura da parte delle strutture nervose vicine, è chiuso da due veli, uno superiore e uno inferiore, che vanno verso l'ilo del cervelletto: il velo midollare superiore (formato anche da fibre provenienti dal cervelletto) e il velo midollare inferiore (fondamentalmente di natura piale con strutture di tipo ependimale). Il velo midollare inferiore caudalmente presenta un foro, il foro di Magendie, attraverso il quale il liquor può passare nello spazio subaracnoideo.
Il pavimento del 4° ventricolo si estende tanto nel bulbo quanto nel ponte. Ha la forma di una losanga e si continua in alto nell'acquedotto mesencefalico e in basso nel canale midollare centrale. Presenta alcuni solchi e rilevatezze che permettono di individuare aree al cui interno sono presenti importanti nuclei del tronco encefalico. È solcato lungo la linea mediana dal solco mediano, lateralmente al quale si descrive un altro solco longitudinale, il solco limitante. Al centro presenta delle fibre, le strie midollari, che dal solco mediano decorrono lateralmente verso i peduncoli cerebellari inferiori. Il solco mediano divide il quarto ventricolo in due triangoli laterali mentre le strie midollari lo dividono in un triangolo superiore e un triangolo inferiore. Nei triangoli laterali la presenza del solco limitante delimita medialmente delle rilevatezze, le eminenze mediali.
Nel triangolo inferiore, nel terzo inferiore dell'eminenza mediale, si nota un triangolo con base in alto e apice in basso detto trigono dell'ipoglosso. Lateralmente un altro triangolo forma il trigono vagale e lateralmente a questo si estende l'area somato-sensitiva o vestibolare.
Nel triangolo superiore, nell'eminenza mediale, nella zona di passaggio bulbo-pontina, è presente il collicolo del faciale.
Mesencefalo
Del mesencefalo si descrive una parte ventrale e una dorsale o tetto.
La parte ventrale del mesencefalo, al disopra del ponte, è formata da due travate di fibre, i peduncoli cerebrali, che provengono dalle cortecce cerebrali. I due peduncoli divergono a V formando una sorta di croce, la crux cerebri. Nello spazio lasciato libero dai due peduncoli, spazio definito sostanza perforata a causa dei fori formati dal passaggio di vasi, oltre all'emergenza dell'oculomotore, compare l'unica superficie esterna del diencefalo (poiché il resto del diencefalo è inglobato nel cervello per via dell'origine embrionale), l'infundibolo che si continua nella neuroipofisi.
Il tetto del mesencefalo è formato da quattro rilevatezze, due per lato, due superiori e due inferiori, i collicoli. I due collicoli superiori appartengono alle vie visive mentre i due collicoli inferiori alla via acustica. Dai collicoli partono delle fibre verso il talamo che formano i bracci dei collicoli, superiore e inferiore.
Emergenze dei nervi encefalici
Le emergenze dei nervi cranici non corrispondono necessariamente alla posizione del nucleo (o dei nuclei) da cui originano.
Dall'alto verso il basso si descrivono nel tronco dell'encefalo 10 delle 12 paia dei nervi cranici (sono infatti esclusi i nervi ottici e olfattivi che non originano dal tronco encefalico) che emergono anteriormente o lateralmente al tronco encefalico, eccetto il nervo trocleare che emerge posteriormente.
Il nervo oculomotore, 3° paio dei nervi cranici, emerge ventralmente nel mesencefalo nello spazio lasciato libero dai peduncoli cerebrali.
Il nervo trocleare, 4° paio dei nervi cranici, emerge nella parte dorsale del tronco encefalico al di sotto dei collicoli inferiori per poi avvolgere lateralmente il trono encefalico e proseguire anteriormente.
Il nervo trigemino, 5° paio dei nervi cranici, emerge con due rami lateralmente sulla parte basilare del ponte. I due rami rappresentano le componenti sensitiva e motoria del trigemino. La componente sensitiva è molto più voluminosa di quella motoria ed è disposta postero-lateralmente rispetto ad essa.
Il nervo abducente, 6° paio dei nervi cranici, emerge nel solco bulbo-pontino tra il ponte e le piramidi.
Il nervo faciale, 7° paio dei nervi cranici, emerge lateralmente nel tronco encefalico, nell'angolo formato dal bulbo, dal ponte e dal cervelletto (angolo ponto-cerebellare), e presenta due radici che sono, la mediale, la radice motoria e la laterale, detta nervo intermedio, che è sensitiva parasimpatica.
Il nervo vestibolo cocleare o acustico, 8° paio dei nervi cranici, emerge lateralmente al nervo faciale, in corrispondenza dei peduncoli cerebellari inferiori.
Il nervo glossofaringeo, 9° paio dei nervi cranici, emerge rostralmente, con diverse radicole, nel solco retroolivare.
Il nervo vago, X paio dei nervi cranici, emerge con più radicole caudalmente al glossofaringeo nel solco retroolivare.
Il nervo accessorio, XI paio dei nervi cranici, ha due componenti, una spinale e una encefalica. La componente encefalica emerge caudalmente al vago nel solco retroolivare. La componente spinale emerge dal midollo spinale e risale per unirsi alla componente encefalica.
Il nervo ipoglosso, XII paio dei nervi cranici, emerge con tante radicole, nel solco preolivare.
Nuclei, fasci e sezioni trasversali del tronco encefalico
In sezione il ponte appare formato da una parte basilare e una parte tegmentale la quale dorsalmente forma il pavimento del quarto ventricolo.
Nel mesencefalo si distinguono ventro-dorsalmente i peduncoli cerebrali, il tegmento e, a chiudere dorsalmente l'acquedotto mesencefalico, il tetto con i tubercoli quadrigemini.
Via dei cordoni posteriori
La via dei cordoni posteriori termina, a livello del bulbo, nei nuclei gracile, cuneato e cuneato laterale. Da qui partono numerose fibre che decussano e si portano al davanti medialmente. Nel decussare formano le fibre arciformi interne che poi si raccolgono lateralmente alla linea mediana, tra il solco mediano e i nuclei olivari, per volgere rostralmente come lemnisco mediale. Nel lemnisco mediale le fibre rappresentano le percezioni provenienti dal soma controlaterale in senso caudo-rostrale in senso ventro-dorsale. Per cui le fibre che portano informazioni provenienti dall'arto inferiore (gracile) si trovano ventralmente alle fibre che portano informazioni dall'arto superiore (cuneato).
Nel ponte il fascio comincia a spostarsi lateralmente e le fibre provenienti dal gracile si spostano lateralmente alle fibre provenienti dal cuneato.
Nel mesencefalo le fibre decorrono lateralmente nel tegmento mesencefalico ed infine terminano nel talamo, nei nuclei ventrali postero-laterali.
Fasci spino-talamici e spino-reticolari
Formano il lemnisco spinale e sono disposti lateralmente nel tronco encefalico. Nel bulbo si trovano dorsalmente alle olive bulbari mentre nel ponte e nel mesencefalo sono disposti lateralmente al lemnisco mediale. I fasci spino-talamici terminano nel talamo nei nuclei ventrali postero-laterali. La via spino-reticolo-talamica termina ai nuclei intralaminari del talamo (i quali proiettano allo striato).
Fasci spino-cerebellari
Il fascio spino-cerebellare dorsale procede verso il cervelletto attraverso i peduncoli cerebellari inferiori omolaterali. Il fascio spino-cerebellare ventrale procede anche nel ponte e attraverso i peduncoli cerebellari superiori decussa e si porta al cervelletto.
Via cortico-spinale
I fasci di fibre provenienti dalla corteccia cerebrale attraversano per intero il tronco encefalico. A livello del mesencefalo contribuiscono a formare i peduncoli cerebrali. Nel ponte decorrono nella parte basilare e nel bulbo formano le piramidi (circa 4 milioni di fibre). Nella parte più caudale del bulbo circa l'80% dei fasci di fibre decussano, formando la decussazione delle piramidi, per formare nel midollo il fascio cortico-spinale. Il 20% delle fibre invece discende omolateralmente.
(Da un nucleo posto ventralmente alle piramidi, nucleo arcuato, nascono le fibre arciformi esterne e altre fibre che si portano dorsalmente a formare le strie midollari.)
Nuclei dei nervi cranici
A livello del collicolo superiore, nel tegmento mesencefalico, inizia, lateralmente, il nucleo mesencefalico del trigemino. Medialmente e più ventralmente comincia a delinearsi il nucleo dell'oculomotore (formato in realtà da diversi nuclei), dorsalmente al quale si pone il nucleo di Edinger-Westphal (parasimpatico). Procedendo caudalmente, ma sempre a livello del collicolo superiore, il nucleo mesencefalico del trigemino continua lateralmente e il nucleo dell'oculomotore medialmente; non compare più il nucleo di Edinger-Westphal.
Caudalmente, a livello del collicolo inferiore, il nucleo mesencefalico del trigemino continua lateralmente mentre medialmente (e caudalmente al nucleo dell'oculomotore) appare il nucleo del trocleare.
Il nucleo mesencefalico del trigemino continua lateralmente nel tegmento mesencefalico e medio-ventralmente ad esso, a livello pontino rostrale, si dispone il locus coeruleus.
A livello del pavimento del 4° ventricolo c'è la presenza di diversi nuclei.
A livello del pavimento pontino, nel tegmento pontino, il nucleo mesencefalico del trigemino continua lateralmente. Ventralmente ad esso inizia il nucleo motore del trigemino e lateralmente il nucleo principale del trigemino. Dorsalmente al nucleo mesencefalico del trigemino si pone il nucleo vestibolare superiore.
Caudalmente, a livello del collicolo del faciale, nel tegmento pontino si distingue dorso-medialmente (ma più lateralmente rispetto alla linea mediana) il nucleo dell'abducente. Più ventralmente e lateralmente c'è il nucleo del faciale, lateralmente al quale continua il nucleo principale del trigemino. Dorso-lateralmente si dispongono i nuclei vestibolari superiore, laterale e mediale.
A livello della zona di passaggio bulbo-pontina si notano dorso-lateralmente i nuclei vestibolari laterale mediale e inferiore e ventralmente ad essi l'inizio della componente spinale del trigemino, il nucleo spinale del trigemino. Ventralmente ad esso e un po' più medialmente continua il nucleo del faciale.
A livello dei recessi del 4° ventricolo, caudalmente al solco bulbo-pontino, lateralmente, nella base dei peduncoli cerebellari inferiori, compaiono i nuclei cocleari, dorsale e ventrale. Medialmente ad essi continuano i nuclei vestibolari mediale e inferiore. Medialmente ai nuclei vestibolari compare il nucleo del tratto solitario costeggiato lateralmente dal tratto solitario. Ventralmente ai nuclei vestibolari prosegue il nucleo spinale del trigemino costeggiato lateralmente dal tratto spinale del trigemino. Ventralmente al nucleo del trigemino compare, tra il nucleo del trigemino e l'oliva bulbare, il nucleo ambiguo. Rostralmente al nucleo ambiguo sono situati i nuclei salivatori superiore e inferiore.
A livello del margine rostrale del nucleo cuneato termina, medialmente ad esso, il nucleo vestibolare inferiore. Medialmente al nucleo vestibolare inferiore continua il nucleo solitario con il suo tratto. Medialmente al nucleo solitario compare il nucleo motore dorsale del vago situato in corrispondenza del trigono vagale e medialmente ad esso il nucleo dell'ipoglosso situato in corrispondenza del trigono dell'ipoglosso. Ventralmente al nucleo cuneato prosegue il nucleo spinale del trigemino ventralmente al quale prosegue il nucleo ambiguo.
A livello del margine inferiore dell'oliva bulbare il nucleo del vago si trova dorsalmente, appena lateralmente alla linea mediana. Lateralmente ad esso prosegue il nucleo solitario e ventralmente il nucleo dell'ipoglosso. Lateralmente, ventralmente al nucleo cuneato, prosegue il tratto spinale del trigemino e ventralmente ad esso il nucleo ambiguo.
Al livello di transizione tra bulbo e midollo, ove le piramidi decussano, il nucleo spinale del trigemino continua nella posizione che nel midollo è occupata dal corno dorsale della sostanza grigia.
Nuclei propri del tronco encefalico
A livello bulbare, dorso-lateralmente alle piramidi si espande in direzione caudo-rostrale quasi fino al solco bulbo-pontino il nucleo olivare inferiore, grosso nucleo di forma a tasca con cavità mediale (ilo) avvolto da un mantello di fibre (amiculum olivae).
A livello pontino, ventralmente al nucleo del faciale, c'è il nucleo olivare superiore. In tutta la parte basilare del ponte sono inoltre presenti nuclei pontini.
Nel mesencefalo dorsalmente ai peduncoli cerebrali si estende la sostanza nera composta da una parte detta pars-compatta e una parte detta pars-reticolata. Medialmente si estende l'area tegmentale ventrale. A livello dei collicoli quadrigemini tra la parte dorsale e la parte ventrale del tegmento si espande un grosso nucleo, il nucleo rosso mentre nel tetto sono presenti i nuclei che formano i collicoli.
Lungo tutto il tronco encefalico sono presenti ulteriori nuclei che formano la formazione reticolare, così nominata perchè è formata da fasci di fibre che formano reti nelle cui maglie sono contenuti i nuclei dei neuroni. L'altro nome per la reticolare è "sistema attivatore ascendente" poiché è responsabile della attivazione della corteccia. I nuclei della formazione reticolare sono suddivisi in nuclei chimicamente definiti e nuclei non chimicamente definiti. Tra i nuclei chimicamente definiti vi sono i nuclei del rafe (serotoninergici) che sono posizionati lungo tutto il tronco encefalico a livello del rafe mediano (zona mediana di incrocio delle fibre del tronco encefalico) e che si dividono in nuclei bulbari, pontini e mesencefalici. Medio-ventralmente al nucleo mesencefalico del trigemino c'è il locus coeruleus (noradrenergico), molto ampio anche se formato da un esiguo numero di cellule. Nella parte più caudale del mesencefalo, nella parte peduncolare del ponte, c'è latero-ventralmente il nucleo peduncolopontino (colinergico). Altri nuclei non chimicamente definiti si estendono lungo tutto il tronco encefalico e sono raggruppati in nuclei dell'area mediana(centrale), nuclei dell'area paramediana gigantocellulari, nuclei dell'area laterale parvicellulare e nuclei precerebellari.
Altri fasci del tronco
Il fascicolo tegmentale centrale è un fascio situato dorso-medialmente ai lemnischi e nel ponte e nel mesencefalo dorsalmente alla decussazione dei peduncoli cerebellari superiori.
Il fascicolo longitudinale mediale decorre vicino alla linea mediana.
A livello pontino ventro-medialmente ai nuclei abducente e faciale le fibre provenienti dai nuclei cocleari nel portarsi controlateralmente formano il corpo trapezoide che incrocia il lemnisco mediale. Le stesse fibre, dopo aver decussato formano il lemnisco laterale che si porta alle olive superiori. Medialmente al lemnisco mediale decorre il lemnisco trigeminale che trasporta fibre dai nuclei sensitivi del trigemino sia omolateralmente che controlateralmente.
Le fibre del nervo ipoglosso dal nucleo omonimo decorrono antero-lateralmente, passano tra i nuclei olivari inferiori e il lemnisco mediale e poi emergono.
Le fibre del nervo faciale decorrono prima dorsalmente, avvolgono il nucleo dell'abducente determinando un ginocchio, il ginocchio interno del faciale che dorsalmente determina il collicolo del faciale, per poi decorrere anteriormente.
Le fibre del trocleare si portano posteriormente e decussano nel tetto del mesencefalo per poi emergere posteriormente.
Le fibre dirette dai nuclei sensitivi trigeminali al talamo ventrale postero-mediale decussano e si uniscono alla via spino-talamica o decorrono medialmente ad essa per quanto riguarda i nuclei spinali e principale del trigemino. Dal nucleo principale nasce anche una via omolaterale che forma il fascio trigemino-talamico posteriore. Tutti i fasci trigemino-talamici sono nel complesso detti lemnisco trigeminale.
Peduncoli cerebellari
I peduncoli cerebellari inferiori contengono fibre olivo-cerebellari, spino-cerebellari dorsali e vestibolari. Originano dal bulbo e a livello pontino entrano nel cervelletto medialmente ai peduncoli cerebellari medi.
I peduncoli cerebellari medi contengono fibre ponto-cerebellari e costituiscono le braccia del ponte (brachia pontis)
I peduncoli cerebellari superiori contengono fibre spino-cerebellari ventrali ma in massima parte fibre dentato-talamiche. A livello mesencefalico decussano formando la decussazione dei peduncoli cerebellari superiori.
Tronco encefalico
Funzioni e correlazioni
Il tronco encefalico è la più grande area di integrazione. Nel il tronco viaggiano, si fermano e vengono integrate informazioni dirette alla corteccia dalla periferia e istruzioni provenienti dalla corteccia verso tutta la periferia. L'organizzazione inframmezzata fibre-nuclei indica che le interazioni in questa zona aumentano notevolmente. Questa interazione raggiunge il massimo nella formazione reticolare. Contribuisce inoltre, soprattutto grazie alla formazione reticolare, alla regolazione dell'attivazione dei centri sovracorticali.
L'area postrema, in corrispondenza dell'obex, è una delle 4 strutture che fanno eccezione alla regola generale del rapporto tra neuroni e letto vascolare che non entrano mai in contatto essendo separati dalla barriera ematoencefalica. Qui i neuroni sono molto resistenti alle variazioni ioniche e sono in diretto contatto con l'albero vascolare. Sono sensori diretti del sangue e loro funzione è quella di essere la zona di innesco del vomito. Il tronco dell'encefalo è la zona che dà vita ai nervi coinvolti (fino al cardias) nell'innesco del vomito per cui sono deputate all'innesco della contrazione antiperistaltica della muscolatura. È una struttura difensiva sensibile alle tossine o forti variazioni ioniche. (Molti antineoplastici inducono l'attivazione di questa zona.)
La via dei cordoni posteriori termina nei nuclei gracile, cuneato e cuneato laterale. I neuroni di questi nuclei proiettano poi al nucleo ventrale postero laterale del talamo attraverso il lemnisco mediale ma la loro funzione non è semplicemente quella di riproiettare al talamo le informazioni propriocettive del tronco e degli arti. Le connessioni che avvengono all'interno dei nuclei permettono di integrare e selezionare una determinata informazione (inibendo la trasmissione delle altre) in base alla sua "forza" rispetto alle altre o in base ad una istruzione della corteccia cerebrale.
I fasci spino-talamici terminano nel nucleo ventrale postero laterale del talamo ma nel tragitto lasciano numerosi collaterali a vari nuclei della formazione reticolare. I fasci spino-reticolari terminano nei nuclei della formazione reticolare.
I fasci spino-cerebellari, ventrale e dorsale, portano le informazioni propriocettive al cervelletto.
I peduncoli cerebrali sono formati da 20 milioni di fibre discendenti dalla corteccia, di cui 16 milioni terminano nel tronco dell'encefalo e i restanti 4 milioni procedono per la via cortico-spinale.
Nei peduncoli cerebrali viaggiano fibre cortico-pontine, che terminano nei nuclei pontini i quali poi proiettano al cervelletto, fibre cortico-bulbari, destinate ai nuclei motori dei nervi cranici, fibre cortico-spinali, che formano poi le piramidi, e fibre che terminano nella formazione reticolare.
Nei peduncoli cerebellari superiori viaggiano informazioni dal cervelletto al nucleo ventrale laterale del talamo.
All'oliva inferiore arrivano fibre dalla corteccia (proiezione cortico-olivare ), fibre provenienti da tutto il midollo spinale (fascio spino-olivare), fibre ponto-olivari dal trigemino e dai nuclei vestibolari. Dall'oliva parte una grande efferenza per il cervelletto. L'oliva penetra nel cervelletto tramite il peduncolo cerebellare inferiore formando il fascio olivo-cerebellare. Queste fibre formano nel cervelletto le fibre rampicanti (che sono solo esclusivamente fibre olivari).
La sostanza nera mesencefalica è così detta perchè i neuroni di quell'area con il passare del tempo diventano scuri, fino a rendere nera l'area, per la presenza di un pigmento che si accumula all'interno delle cellule, la neuromelanina, che non ha nulla a che vedere con la melanina (infatti la neuromelanina è presente anche negli albini, ove invece manca la melanina).
La sostanza nera non è una struttura unica. Una parte, più craniale, è caratterizzata da cellule molto vicine tra loro che formano la pars-compacta della sostanza nera. L'altra parte è formata da cellule più distanti tra loro, formanti un reticolato, che formano la pars-reticulata della sostanza nera. Queste due componenti sono diverse, sia per aspetto che per funzioni.
Le cellule che formano la pars-compatta della sostanza nera sono neuroni dopaminergici identificabili come tali in immagine immunoistochimiche in cui sono usati come marker anticorpi anti-tirosina idrossilasi (enzima chiave nella produzione di dopamina). I neuroni della pars-compatta proiettano al cervello e precisamente ai nuclei della base (o gangli della base) formando la proiezione nigro-striata. La morte delle cellule dopaminergiche della sostanza nera pars-compatta e la conseguente scomparsa della proiezione nigro-striatale sono alla base del morbo di Parkinson. Quindi, poichè nel morbo di Parkinson c'è la scomparsa delle cellule dopaminergiche, se ne deduce che la proiezione nigrio-striatale è centrale nella motricità, almeno in determinati aspetti della motricità diversi dall'efficacia e dall'efficienza (efficacia ed efficienza sono sotto il controllo del cervelletto).
La sostanza nera pars-reticulata è formata da neuroni GABAergici e sono un'estensione mesencefalica del pallido interno (nucleo dello striato).
Nella parte mediale più ventrale del tegmento mesencefalico c'è poi l'area tegmentale ventrale, anch'essa formata da cellule dopaminergiche simili a quelle della sostanza nera pars compatta, che proietta in massima parte alla zona più ventrale del corpo striato (nucleo accumbens) formando la proiezione meso-accumbale e in parte alla corteccia frontale che gira intorno al corpo calloso (corteccia del cingolo) ovvero al sistema limbico (proiezione meso-cortico-limbica). Poiché il nucleo accumbens è il centro edonistico per eccellenza nel cervello la proiezione meso-cortico-limbica risulta essenziale nello svolgere azioni che richiedono gratificazione. Ogni essere vivente non fa nessuna azione se non in vista di un premio o di una gratificazione. Anche se in alcuni casi può sembrare che non ci siano premi va considerato che il primo premio è la non punizione. Studi sull'apprendimento degli animali da laboratorio dimostrano che si ottengono più risultati usando come "premio" la non punizione che una gratificazione. Il circuito meso-accumbale è centrale in tutte le aspettative premianti che sono finalizzate al mantenimento di se stessi e della specie (assunzione di cibo, di liquidi e attività sessuale).
il neurone dopaminergico è centrale, ha una azione nei confronti del nucleo accumbens. il nucleo accumbens di norma è inibitorio, ha azione inibitoria nella proiezione strio accumbo nigrale (perchè questo di norma viene mantenuto inibto). la presenza di stimoli sensoriali va a inibire l'interneurone che controlla il neurone dopaminergico, neurone dopaminergico che sono per lo piu mantenuti inibiti. poi danno un grande colpo e stimola il nucleo accumbens
La presenza di stimoli sensoriali inibisce l'interneurone che controlla il neurone dopaminergico del nucleo accumbens.
Il nucleo accumbens proietta con le stesse modalità dello striato.
Qui subentrano anche le scelte perchè permette di scegliere tra due alternative quella che darà più piacere.
Questo è anche il circuito delle dipendenze. Sia il nucleo accumbens che l'area tegmentale ventrale presentano recettori per gli oppioidi (endogeni, ma anche esogeni come endorfine, morfina, eroina) e per l'alcol (o forse sono particolarmente influenzati dall'alcool). I residui alcolici sono in grado di inibire l'interneurone del nucleo accumbens con conseguente attivazione (disinibizione) del nucleo accumbens. Il nucleo accumbens è talmente condizionante che se ad un topino si infila un elettrodo che stimola il nucleo accumbens e gli si fa provare che spingendo una leva si ha la stimolazione del nucleo accumbens, il topo non si sposta di li finchè non muore (non si sposta nemmeno per mangiare e bere).
L'effetto della cocaina e degli anfetamino-simili è diverso. Non si ha diretta stimolazione degli elementi accumbali o dell'area tegmentale ventrale ma l'azione è sulla membrana del neurone dopaminergico. Il terminale assonico presinaptico del neurone dopaminergico rilascia dopamina. La dopamina dopo un certo tempo viene ricaptata dallo stesso terminale presinaptico (ad opera di un trasportatore transmembrana della dopamina) che determina l'end point della trasmissione. La cocaina e gli anfetamino-simili bloccano il trasportatore della dopamina (sono falsi ligandi) per cui la trasmissione dopaminergica non viene interrotta. C'è quindi over-espressione della dopamina e l'effetto sulla persona è simile ai sintomi di un tourettico.
I nuclei della formazione reticolare hanno diverse afferenze e diverse efferenze.
I nuclei del rafe fanno parte dei nuclei chimicamente definiti in quanto producono serotonina (nuclei serotoninergici)e la serotonina è il neurotrasmettitore che inibisce il dolore.
I nuclei del bulbo, che formano il nucleo oscuro, il nucleo magno e il nucleo parvicellulare del rafe, danno vita alla via rafe-spinale che nel midollo spinale la ha come effetto finale l'inibizione della trasmissione della nocicezione.
I nuclei pontini del rafe proiettano al cervelletto entrando a far parte delle fibre muscoidi.
I nuclei mesencefalici del rafe proiettano alla corteccia, allo striato e a gran parte del talamo. La proiezione rafe-corticale forma una sinapsi asso-assonica con un terminale assonico talamico, diretto ad un dendrite della corteccia (la proiezione talamo-corticale, dal talamo alla corteccia, è glutammatergica), che possiede recettori per la serotonina la quale provoca ulteriore rilascio di glutammato (azione facilitatoria della serotonina). Il prozac, un serotonino-mimetico, mimando l'azione della serotonina svolge la sua azione antidepressiva (a riprova dell'azione anti-tristezza della serotonina, anche se è l'aumento conseguente della proiezione glutammatergica ad avere azione antidepressiva). Quindi anche in questo caso la proiezione serotoninergica è "antidolorifica"(dolore mentale, poiché il cervello può trovarsi in diversi stati tra cui la tristezza).
Il locus coeruleus, altro nucleo chimicamente definito contiene neuroni noradrenergici, pochi neuroni ma con un ampia proiezione: midollo spinale, cervelletto, diencefalo, corteccia cerebrale (in due strati). L'ampia distribuzione del sistema noradrenergico indica che esso non è selettivo come gli altri sistemi chimicamente definiti (per esempio la proiezione circoscritta dei neuroni dopaminergici indica una loro funzione specifica nella regolazione di determinati stati specifici della mente) e controlla stati generali del cervello (propri della formazione reticolare) come il ritmo sonno-veglia.
L'ultimo sistema chimicamente definito usa l'acetilcolina come neurotrasmettitore ed è particolarmente rappresentato dal nucleo peduncolo pontino. L'innervazione colinergica da questo nucleo va in parte al cervelletto, in parte al midollo spinale ma in massima parte al talamo. Questa innervazione rientra nei sistemi dello stato di veglia. L'innervazione colinergica della corteccia non nasce dal tronco ma da due piccoli nuclei basali, il nucleo basale che sta alla base dei gangli della base e dai nuclei mediali del setto. I neuroni di questi nuclei muoiono nel morbo di Alzheimer (ma non ne sono la causa) in cui.il soggetto è disorientato e ha perdita della memoria (sbaglia la via di casa e/o non riconosce moglie e/o figli) Poiche nel morbo di Alzheimer viene a mancare l'innervazione colinergica dell'encefalo, della corteccia e dell'ippocampo a posteriori si può dire che l'innervazione colinergica della corteccia e dell'ippocampo è centrale nell'orientamento temporo-spaziale e nella memoria (specie quella a breve termine).
Il passaggio dallo stato di veglia allo stato di sonno è gestito dai sistemi della formazione reticolare, sia quelli chimicamente definiti sia quelli non chimicamente definiti. Per cui il passaggio tra veglia e sonno non sono proprietà della corteccia ma del tronco. (La veglia è un range continuo ma ci sono diversi stadi come la fase di attenzione, e l'attenzione può essere focalizzata fino all'allerta.)
Le cellule del locus coeruleus, avendo una così ampia proiezione, funzionano quasi all'unisono e infatti hanno fra loro sinapsi elettriche (gap junction). L'innervazione noradrenergica, insieme a quella serotoninergica, gioca un ruolo fondamentale nel passaggio dallo stato di veglia a quello di sonno.
Nello stato di sonno riconosciamo 2 grandi fasi:
-il sonno non REM (Rapid Eye Mouvement, condizione in cui il soggetto ha una separazione tra il talamo e la corteccia e attiva rapidi movimenti oculari mentre dal punto di vista muscolare è completamente disteso)
-il sonno REM durante il quale la corteccia, liberata dalle afferenze corporee, crea il sogno che rappresenta uno stato di coscienza vivido.
Un danno alla formazione reticolare è la causa dello stato di coma. Un soggetto, per svariate cause, può avere un danno di grado grave, medio, moderato, lieve o elevato alla formazione reticolare. Il soggetto passa in questi casi da uno stato di coscienza ad uno stato di incoscienza. Il soggetto che va in coma, presenta danneggiamenti al tronco dell'encefalo. Dal coma o si muore o si esce. Quando si esce si possono avere diversi deficit dello stato di coscienza. Il più grave è quando si ha lo stato vegetativo persistente. Poi c'è lo stato vegetativo temporaneo che dura da 4 a 6 mesi dal quale si esce. In alcuni casi il paziente può rispondere (ad esempio con il movimento degli occhi) ad alcuni stimoli. In altri casi il paziente è completamente cosciente ma è locked in, chiuso dentro. Il paziente non riesce a esprimere ciò che pensa.
I nuclei reticolari precerebellari proiettano, come fibre muscoidi, al cervelletto perchè anche il cervelletto deve avere la sua attivazione dalla formazione reticolare.
I neuroni gigantocellulari dell'area paramediana sono caratterizzati dal fatto di avere un assone ascendente verso la corteccia, le strutture talamiche ecc, o discendente verso il midollo spinale. Altri assoni si dividono e danno sia rami ascendenti che discendenti tanto per la corteccia che per il midollo spinale. In ogni caso la collateralizzazione di questi assoni è notevolissima e ogni neurone è legato a tantissimi altri neuroni e ciò fa si che questa formazione funzioni come un tutt'uno. L'area paramediana è sostanzialmente punto di efferenze.
L'area parvicellulare è l'area delle afferenze. Presenta un centro espiratore la cui stimolazione può provocare l'accelerazione del battito cardiaco e l'innalzamento della pressione arteriosa.
Superiormente all'area parvicellulare c'è l'area parabrachiale che presenta numerose connessioni. Le fibre afferenti provengono dal nucleo del tratto solitario, dalla corteccia dell'insula e da parti adiacenti al lobo parietale. Le efferenze proiettano all'ipotalamo, all'area preottica, ai nuclei intralaminari e all'amigdala. Può includere anche un "centro pneumotassico" riconosciuto dai fisiologi come regione implicata nel controllo del ritmo della respirazione. Lesioni di questa regione possono provocare respirazione apneusica nella quale si verifica una pausa di qualche secondo tra una inspirazione profonda e l'inizio dell'espirazione.
In generale la formazione reticolare riceve informazioni da:
-la corteccia prefrontale
-informazioni di tipo vegetative dall'ipotalamo
-proiezioni dal cervelletto
-dal midollo
-dai nuclei dei nervi encefalici e dall'apparato vestibolare.
E tramiti i vasti collegamenti dei suoi svariati nuclei manda efferenze di diverso tipo a tutto il SNC.
Funzioni e correlazioni dei nuclei dei nervi cranici e dei nervi cranici
La grande differenza tra nervi spinali e cranici è che mentre gli spinali sono quadrimodali (o somato/viscero-motori o somato/viscero-sensitivi) i cranici hanno ognuno una modalità proprie.
Le emergenze dei nervi sono origini apparenti perchè i nuclei dei nervi possono stare anche in altri luoghi diversi dalle emergenze (come ad esempio il nucleo dell'ipoglosso che sta dorsalmente).
Per alcuni nervi cranici si parlare anche di viscerale "speciale" termine che si riferisce ai nervi destinati alle strutture che originano dagli archi e dalle tasche branchiali. Tutte queste strutture (come i muscoli mimici o la laringe) non sono soma e non sono visceri e sono sottoposte a un duplice controllo, uno automatico e uno volontario.
Alla decussazione della piramidi si nota una diversa disposizione dei nuclei rispetto al midollo spinale. Nel midollo in direzione ventro-dorsale ci sono i nuclei somatomotori, i visceromotori(nel toracico), quindi strutture viscero sensitive e somato sensitive. Qui, data la mancata chiusura del tetto, ciò che era ventrale è diventato (all'incirca) mediale e ciò che era dorsale è diventato (all'incirca) laterale.
Per questo motivo i nuclei sono schematizzati lungo dei piani in senso medio-laterale e rostro-caudale detti colonne nucleari. Medialmente ci sono le colonne motrici e lateralmente le colonne sensitivo-sensoriali.
Lateralmente sono riportate quattro colonne afferenti sensitive-sensoriali mentre medialmente solo tre colonne efferenti motorie (manca un efferenza che in realtà non manca).
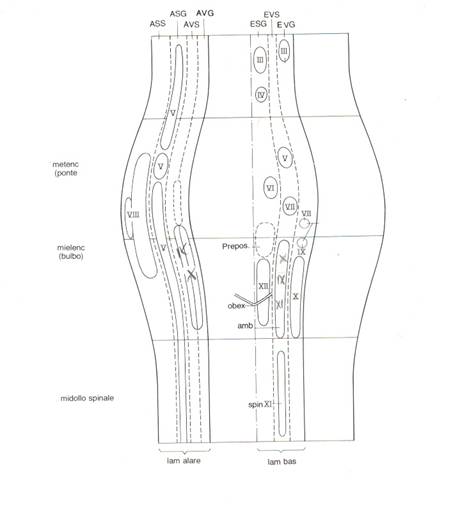
Tra
le colonne efferenti la più mediale riporta le efferenze somatiche generali
ovvero i nuclei motori destinati a strutture somatiche generali. Tale colonna
presenta la V dell'obex e a cavallo dell'obex
il nucleo dell'ipoglosso. Risalendo nella colonna si trovano il 6° a
livello pontino, il 4° e il 3° (il 3° non è un nucleo unico ma ogni muscolo ha
il proprio nucleo) a livello mesencefalico destinati alla motricità dei muscoli
estrinseci dell'occhio.
Lateralmente compare la colonna delle efferenze viscerali speciali. Il primo nucleo,a livello bulbare, contiene più nuclei che danno fibre al vago (nucleo ventrale del vago) al glossofaringeo e all'accessorio ed è perciò detto nucleo ambiguo. Tramite i detti nervi il nucleo ambiguo innerva il faringe e il terzo craniale dell'esofago (che ha una muscolatura di tipo striato sottoposta al controllo volontario come anche il costrittore inferiore del faringe), i muscoli del cavo orale e del laringe. Poi, a livello pontino, c'è il nucleo del faciale che innerva muscoli mimici, un ventre del digastrico e lo stapedio e superiormente il nucleo motore del trigemino.
Lateralmente è disposta la colonna delle efferenze viscerali generali. Il nucleo maggiore, a livello bulbare, è il nucleo del X, nucleo parasimpatico, detto nucleo motore dorsale del vago o centro cardio-pneumo-enterico perchè il vago, come efferenza parasimpatica, arriva fino al colon sigma.
Rostralmente ci sono i nuclei parasimpatici salivatori, salivatorio inferiore, bulbare, e salivatorio superiore, pontino. Il salivatorio inferiore manda le sue fibre con il 9° paio dei nervi cranici ed innerva la parotide. Il salivatorio superiore va con il 7° ed innerva la sottolinguale e sottomandibolare. L'innervazione parasimpatica del nucleo salivatorio superiore va anche alla ghiandola lacrimale[1].
In cima alla colonna, a livello mesencefalico, c'è il nucleo parasimpatico del 3°, il nucleo di Edinger-Westphal (fibre pregangliari colinergiche).
Tra le colonne sensitive la più laterale è quella delle afferenze somatiche speciali provenienti dall'orecchio e all'interno di questa colonna giunge il nervo vestibolo-cocleare che non essendo un nervo unico termina in due nuclei: uno più piccolo laterale e uno 4 volte più grosso mediale. Il nervo cocleare afferisce al più piccolo nei due nuclei cocleari, il cocleare ventrale e dorsale. Il vestibolare termina nei 4 nuclei vestibolari. Medialmente c'è la colonna delle afferenze somatiche generali in cui sono presenti tutti i nuclei del trigemino (dallo spinale al mesencefalico). Nelle ultime due colonne arrivano le afferenze viscerali generali e speciali accolte dal nucleo del tratto solitario. I maggiori tributari delle afferenze viscerali generali sono il vago e il glossofaringeo.
In direzione caudo-rostrale il primo nervo cranico è il 12°, l'ipoglosso, che origina da una serie di radicole lungo il solco preolivare. Esso è monomodale poichè è solo somatomotore e innerva tutti i muscoli della lingua. Riceve, oltre alle afferenze corticali, anche afferenze dai nuclei del tratto solitario e sensitivi del trigemino potendo così integrare tali informazioni e determinare adeguate risposte riflesse necessarie alla corretta suzione, masticazione e deglutizione.
I nervi accessorio, vago e glossofaringeo (11°, 10° e 9° paio) sono nervi con diverse modalità ed origini.
Il nervo accessorio ha un origine spinale (rami emergenti da C3 C2 e C1 che risalgono lungo il midollo), che forma la componente somatomotrice del nervo, che si unisce poi con la componente bulbare originata dal nucleo ambiguo.
Le due componenti procedono come nervo accessorio fino al foro giugulare perchè dopo le fibre del nervo accessorio che originano dal nucleo ambiguo lasciano il nervo accessorio e procedono con il vago. A questo punto il nervo accessorio, composto di sole fibre spinali, si porta ad innervare i muscoli sternocleidomastoideo e trapezio.
Il nervo vago ha 6 modalità: è visceromotore, viscerosensitivo, somatomotore, somatosensitivo, sensitivo speciale.
Le fibre gustative e viscerosensitive terminano nel nucleo del tratto solitario.
Le fibre somatosensitive terminano nel nucleo spinale del trigemino.
Le fibre somatomotrici originano dal nucleo ambiguo mentre le fibre visceromotrici parasimpatiche originano dal nucleo motore dorsale del vago e in parte anche dal nucleo ambiguo.
I corpi cellulari delle fibre gustative del vago sono situati nel ganglio inferiore (nodoso) del nervo vago. Le fibre gustative innervano insieme alle fibre del glossofaringeo il terzo posteriore della lingua, la faringe e in più i calici gustativi dell'epiglottide. Tali fibre terminano nel nucleo solitario nella sua porzione rostrale, detta anche nucleo gustativo.
I corpi cellulari delle fibre che portano la sensibilità viscerale generale sono anch'esse situati nel ganglio nodoso e le fibre, che portano la baro e chemocezione dall'arco aortico oltre agli impulsi sensitivi viscerali dei visceri del torace e dell'addome, terminano nella parte caudale del nucleo solitario.
I corpi cellulari delle fibre somatosensitive sono situati nel ganglio superiore (giugulare) del vago. Perifericamente innervano la parte inferiore della faringe, l'esofago, la laringe, l'albero tracheo-bronchiale, l'orecchio esterno, la membrana timpanica e (insieme al trigemino) la dura meninge (soprattutto per la componente dolorifera).
Le fibre somatosensitive entrano nel nucleo spinale del trigemino.
Le fibre somatomotrici originate dal nucleo ambiguo innervano tutti i muscoli della faringe (escluso lo stilo-faringeo innervato dal glossofaringeo), dell'esofago e della laringe. Di queste, le fibre che fanno da subito parte del vago innervano i muscoli suddetti esclusi i muscoli intrinseci della laringe e i muscoli del palato molle. Questi ultimi (escluso il tensore del velo palatino, innervato dal trigemino) sono innervati dalle fibre che in origine avevano viaggiato insieme all'accessorio.
Le fibre parasimpatiche pregangliari originate dal nucleo motore dorsale del vago e dal nucleo ambiguo si portano ai visceri toracici (cuore, polmoni ecc) e addominali fino al colon sigma.
Il nervo glossofaringeo, come il vago, ha diverse origini.
Le fibre che formano le afferenze gustative e viscerosensitive terminano nel nucleo del tratto solitario.
Come per il vago, le afferenze somatosensitive terminano nel nucleo spinale del trigemino.
Come il vago, il glossofaringeo origina le fibre somatomotrici dal nucleo ambiguo.
Attraverso il glossofaringeo fibre parasimpatiche originate nel nucleo salivatorio inferiore si portano al ganglio otico come fibre pregangliari (attraverso il nervo piccolo petroso superficiale) e da qui fibre postgangliari vanno alla parotide.
I corpi cellulari delle fibre afferenti sia somatosensitive che viscerosensitive gustative del nervo glossofaringeo sono ubicati nei gangli superiore e inferiore del glossofaringeo mentre i corpi delle fibre viscerosensitive generali sono situati nel ganglio inferiore.
Le fibre viscerosensitive gustative portano insieme a fibre vagali il gusto dal terzo posteriore della lingua e dalla faringe e terminano nel nucleo gustativo del nucleo solitario.
Le fibre viscerosensitive generali si portano perifericamente al seno e al glomo carotico per la baro e chemo-cezione e nel nucleo solitario si dispongono caudalmente.
Le fibre somatosensitive, che terminano nel nucleo spinale del trigemino, si distribuiscono al terzo posteriore della lingua, alla parte superiore della faringe, alla tuba uditiva e all'orecchio medio.
Le fibre somatomotrici sono poche e si portano alla faringe per innervare il muscolo stilo-faringeo.
Le informazioni che giungono al nucleo solitario sono informazioni di tipo viscerali ma diverse da quelle che arrivano dal midollo spinale (perchè nel midollo si tende ad un'integrazione tra soma e visceri contenuti nel soma).
Questa informazione è correlata alla regolazione dei visceri. Infatti dal nucleo solitario partono proiezioni all'ipotalamo nonché alla formazione reticolare.
Medialmente al nucleo del tratto solitario c'è il nucleo motore dorsale del vago, e lateralmente il nucleo ambiguo e ad essi giungono proiezioni dall'ipotalamo, dalla formazione reticolare oltre che direttamente dal nucleo solitario e dal sistema olfattivo.
Al nucleo ambiguo, inoltre, giungono proiezioni dai nuclei sensitivi del tronco compreso il nucleo spinale del trigemino. A questo livello pertanto si crea un arco riflesso viscerale per l'immediata risposta viscerale ai fini del mantenimento dell'omeostasi (es. Organo recettoriale del seno aortico che contiene pressocettori e chemocettori. Se la pressione, o l'ipercapnia, aumenta c'è di risposta una veloce modificazione della gittata sistolica tramite il nucleo motore dorsale del vago). In questa zona inoltre c'è anche l'area postrema, zona d'innesco del vomito.
Il nervo vestibolo-cocleare va considerato con la via acustica.
Il nervo faciale, il 7°, è un nervo che è unito al 7° bis (intermedio) ed ha una funzione visceromotrice speciale in quanto innerva i muscoli mimici, una funzione viscerosensitiva speciale, una funzione parasimpatica e in parte una funzione somatosensitiva.
Le fibre motorie originano dal nucleo motore del faciale e innervano i muscoli mimici, il ventre posteriore del digastrico, lo stapedio e lo stiloioideo. Al nucleo del faciale giungono diverse proiezioni cerebrali sia per il controllo volontario che per il controllo automatico emotivo della mimica facciale. Il nucleo del faciale riceve inoltre afferenze acustiche dal nucleo olivare superiore per il riflesso del muscolo stapedio.
Le fibre somatosensitive contribuiscono a innervare l'orecchio esterno e hanno i corpi cellulari nel ganglio genicolato. Terminano nel nucleo spinale del trigemino.
Le fibre gustative presentano anch'esse il corpo cellulare nel ganglio genicolato. Dal ganglio genicolato tali fibre in parte procedono nel nervo grande petroso, fuoriescono nella fossa pterigopalatina e contestualmente al mascellare si distribuiscono al palato ove innervano i calici gustativi principalmente sul palato molle.
Altre fibre gustative procedono nel nervo corda del timpano, fuoriescono nella fossa infratemporale e contestualmente al ramo linguale del mandibolare si distribuiscono ai calici dei due terzi anteriori della lingua.
Tutte queste fibre, dal ganglio genicolato al bulbo, formano il nervo intermedio che termina nel nucleo gustativo del nucleo solitario. Da tale nucleo, tra le altre proiezioni, nasce la via gustativa che si porta al talamo ventrale posteriore e da qui alla corteccia.
La componente viscerale generale è parasimpatica e presiede all'innervazione delle ghiandole salivari, sottolinguali e mandibolari, e della ghiandola lacrimale.
Le fibre pregangliari partono dal nucleo salivatorio superiore e nel ganglio genicolato parte procede, attraverso il nervo grande petroso, verso il ganglio pterigopalatino da cui le fibre postgangliari si portano alla ghiandola lacrimale e alle ghiandole della mucosa nasale.
Le fibre restanti sono disposte nel nervo corda del timpano e terminano nel ganglio sottomandibolare da cui le fibre postgangliari innervano le ghiandole sottolinguali e sottomandibolare.
I nuclei salivatori ricevono proiezioni dall'ipotalamo, dalla formazione reticolare dei nuclei solitari e trigeminali spinali. Tali connessioni influenzano la salivazione e la lacrimazione in risposta a stimoli sensitivo-sensoriali oltre che a stati emotivi. (In caso di lunga anestesia o di grosse resezioni intestinali si può avere un'ipogenusia o un'agenusia con perdita parziale o totale del gusto.)
Il 5°, il trigemino, è il nervo più "robusto" e presenta due radici: una grossa radice sensitiva e una piccola radice motoria.
La sensitiva ha fibre il cui corpo è situato nel ganglio di Gasser da cui partono le tre branche del trigemino.
Le fibre sensitive innervano la parte anteriore della testa, la mucosa orale e nasale, i denti, la dura meninge e le arterie cerebrali. Le fibre sensitive terminano nei nuclei pontini (principali) e spinali del trigemino (in generale le sensazioni dolorifiche, termiche e di tatto lieve terminano caudalmente mentre le altre rostralmente). Dai nuclei sensitivi parte la proiezione controlaterale, tramite il lemnisco trigeminale, al talamo ventrale postero-mediale.
Dal nucleo principale del trigemino c'è un'afferenza che è omolaterale al talamo per motivi di simmetria tra destra e sinistra.
Quindi nel nucleo ventrale posteriore del talamo viene ricostruito tutto il corpo nella sua unitarietà. Il talamo proietta poi alla corteccia.
Per quanto riguarda la propriocezione della zona relativa ai muscoli masticatori e della pressione sui denti, i corpi cellulari sono ubicati nel nucleo mesencefalico del trigemino (stando eccezionalmente all'interno del sistema nervoso centrale).
Da questi parte una proiezione per il nucleo motore del trigemino consentendo vari riflessi per la masticazione.
Le fibre motorie partono dal nucleo motore del trigemino e si distribuiscono, tramite un ramo che si unisce alla branca mandibolare, ai muscoli masticatori.
Il trigemino innerva inoltre anche il tensore del velo palatino, il tensore del timpano, il ventre anteriore del digastrico e il miloioideo.
L'innervazione di qualsiasi organo presenta un rapporto duplice perchè l'organo bersaglio si mantiene in vita finchè viene innervato. La neurotrasmissione è un sistema che attiva una segnalazione che è indice di trofismo. La maggior parte dei muscoli infatti quando vengono denervati vanno incontro ad ipotrofia. Per il nervo vale la stessa cosa. Per es. Nella parotide viene prodotto NGF che serve al glossofaringeo per mantenerlo in vita. Se si stacca il glossofaringeo dalla ghiandola c'è una degenerazione dell'assone.
|
|