|
| Appunti superiori |
|
|
| Appunti superiori |
|
| Visite: 4378 | Gradito: |
Leggi anche appunti:Trasformazione fisica e chimica (miscuglio e composto)TRASFORMAZIONE FISICA E CHIMICA (MISCUGLIO E COMPOSTO) OBIETTIVO:Verificare I modelli di trasmissione ereditariaI modelli di trasmissione ereditaria Gli alleli e i fenotipi. I geni sono ColeraCOLERA INCUBAZIONE: media di 5 giorni. SINTOMI: enterite acutissima |
 |
 |
IL CICLO CELLULARE E LE TECNICHE DI SINCRONIZZAZIONE
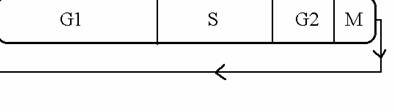
Il ciclo vitale delle cellule si svolge attraverso una serie di eventi metabolici, come sintesi di proteine, di RNA e DNA che con una ben precisa sequenza temporale scandiscono la progressione di una cellula da mitosi a mitosi. Sono proprio questi eventi che controllano la transizione della cellula da una fase all'altra del ciclo cellulare (vedi fig.1).
La cellula neoformata rimane per un certo periodo in uno stato in cui non duplica il suo DNA, ma aumenta la sua biomassa. Questa fase e' detta fase G1, e il contenuto in DNA viene indicato come 2n cioe' come composto dal normale corredo cromosomico (2 n perche' sappiamo che le cellule, a meno di gravi patologie, hanno un certo numero di coppie di cromosomi, numero che e' specifico per ciascuna specie). Quando le cellule si dividono, esse devono duplicare il loro DNA. Questa fase e' di solito preceduta da una fase di intensa sintesi proteica, dato che le cellule devono preparare tutto il complesso macchinario enzimatico che servira' loro alla duplicazione del DNA, nonche' provvedere a duplicare tutti gli organelli, dato che le cellule figlie devono essere egualmente equipaggiate (questo almeno nelle cellule che si dividono in modo simmetrico). La fase di sintesi del DNA viene indicata come fase S, ed ha una durata di tempo caratteristica per ciascun tipo di cellula. Le cellule umane, ad esempio, hanno una durata della fase S che e' di circa 7-9 ore. La divisione cellulare (cioe' la mitosi) occupa un periodo di tempo molto limitato, di circa 0.5-1 ora, ed e' preceduta dalla fase in cui la cellula rimane con un contenuto doppio di DNA (4 n) prima della divisione vera e propria. Questa fase viene indicata come fase G2, mentre la mitosi vera e propria come fase M. Una parte di cellule puo' uscire dal ciclo definitivamente per morte o per differenziamento, oppure temporaneamente entrando nello stato di quiescenza.
|
Figura 1 |
Le cellule in coltura di solito crescono in modo asincrono , cioe' in ogni istante noi troviamo cellule nelle diverse fasi del ciclo. Se la coltura e' in fase esponenziale ci aspettiamo di trovare costante la percentuale di cellule nelle 3 fasi principali (G0/G!, S e G2/M). A confluenza le cellule si accumuleranno nella fase G0/G1, totalmente se e' una coltura di cellule normali, parzialmente se trasformata.
E' possibile sincronizzare le cellule, bloccarle cioe' tutte (o quasi tutte) in uno stadio specifico del ciclo, per poi rilasciarle: le cellule a questo punto dovrebbero muoversi tutte insieme attraverso le varie fasi del ciclo cellulare. Questo puo' essere fatto con diversi sistemi: quelli chimici hanno il grande svantaggio che interferiscono pesantemente con la normale 'vita' cellulare, mentre i metodi che selezionano le cellule in base ad alcune proprieta' fisiche, come ad esempio il minor ancoraggio delle cellule mitotiche o la variazione delle dimensioni e della densita' delle cellule nelle varie fasi del ciclo, sono invece dei sistemi che perturbano meno la popolazione cellulare. I sistemi chimici di solito forniscono una resa maggiore di cellule in una data fase del ciclo. Il metodo da scegliere quindi dipende (come sempre) da quello che si vuole ottenere.
Purtroppo la durata della sincronia e' brevissima, venendo persa spesso gia' dopo un solo ciclo.
1 Selezione di cellule mitotiche.
Shaking: Uno dei metodi piu' semplici per le cellule adese e' la selezione delle cellule mitotiche per scuotimento (shaking) della fiasca. Le cellule devono essere nella fase esponenziale e nelle condizioni ottimali di divisione. Le cellule mitotiche diventano rotonde e poco attaccate, e si riescono a staccare con (relativa) facilita'. La procedura ottimale consiste nell'utilizzare un sistema meccanico per lo scuotimento della fiasca che produca circa 20 colpi in 3 sec, e nel ripetere lo scuotimento e la raccolta delle cellule distaccate ogni 15 minuti per circa 1 ora. Le cellule presenti inizialmente devono essere almeno 107. La resa e' comunque piuttosto bassa (circa il 3-5 %), anche perche' le percentuale di cellule mitotiche di solito e' piuttosto bassa in una popolazione in crescita asincrona. Con lo scuotimento manuale di solito non si supera l'1-3%. La resa puo' essere aumentata trattando per tempi brevi le cellule con colcemide o nocodazolo, e utilizzando comunque un doppio sistema di blocco.
La combinazione di un blocco singolo con timidina (vedi paragrafo 8) per 15-16 ore seguito da 4-5 ore in presenza di nocodazolo 0.04 g/ml, porta ad una resa del 25-30% di cellule mitotiche per shaking, e altre cellule mitotiche si otterranno dagli shaking successivi.
Un protocollo da noi utilizzato sulla linea murina C108 (sarcoma di Lewis) prevede per il blocco in fase M la deprivazione di siero per 24 ore e quindi il trattamento per 18 ore con nocodazolo 660 nM. La stock di nocodazolo (2000 X) viene preparata in DMSO:H2O 1:1. Le cellule mitotiche vengono raccolte e lavate 2 volte con PBS, e quindi rimesse in coltura.
Tripsinizzazione: Alcuni autori (Stubblefield et al., J. cell Physiol., 69, 354, 1967), hanno riportato una metodica per rilasciare le cellule metafasiche mediante trattamento con tripsina diluita (allo 0.1%), utilizzando fibloblasti di criceto cinese DON-C. Questa metodica prevede che le cellule siano coltivate in medium McCoy's 5a addizionato di 20% di FCS e 0.08% di idrolizzato di lactalbumina, e divise ogni 24 ore. Vengono seminate ad una densita'di 1.2 x 105 cell/ml (3 x 104 cell/cm2). Dopo 18 ore dalla semina vengono aggiunti 0.06 g/ml di colcemide e lasciati 4 ore. Alcune cellule mitotiche si staccano e saranno unite a quelle raccolte successivamente. Si rimuove il terreno e lo si sostituisce con tripsina fredda allo 0.1% in PBS. Si scuote (2 cicli/sec) e dopo 45 sec si trasferiscono le cellule che si sono sospese (dovrebbero essere tutte cellule mitotiche) in un tubo da centrifuga e si centrifugano a 400 g x 2 min, risospendendole quindi in medium fresco ad una densita' di 4 x 104 cell/ml. Dopo 20-30 min le cellule si dividono in modo sincrono.
Selezione da microcarrier: Quando le cellule sono cresciute su microcarrier, e' possibile staccare le cellule mitotiche semplicemente aumentando la velocita' di agitazione del sistema.
3 Uccisione selettiva di cellule in particolari fasi.
Viene fatta di solito o utilizzando timidina triziata ad alta attivita' specifica, o con idrossiurea (quest'ultima metodica applicabile solo con cellule suscettibili). Non e' una metodica molto efficace, perche' finisce con il selezionare le cellule che crescono lentamente, e finisce che comunque queste perdono la sincronia velocemente.
4 Selezione delle cellule in base alle dimensioni.
Si basa ovviamente sul principio che le cellule in G1 sono piu' piccole delle cellule in G2. La selezione puo' essere effettuata con strumentazioni particolari, come i citofluorimetri a flusso equipaggiati per il cell sorting, o una particolare tecnica centrifugativa, l'elutriazione. Un certo grado di sincronizzazione si puo' anche raggiungere separando le cellule per centrifugazione su gradiente di densita'.
5 Sincronizzazione per divisione.
E' difficile non introdurre un certo grado di sincronizzazione con le normali operazioni di routine. Sempre Stubblefield ha riportato che la subcoltura frequente delle cellule DON-C (ogni 24 ore) esercita una selezione favorendo le cellule con un tempo di generazione di 12 ore e che la frequenza mitotica e' piu' alta a 7 e a 19 ore dopo lo splitting.
Spesso una coltura cellulare lascia la fase logaritmica ed entra nella fase stazionaria prima di essere divisa. In questo caso e' possibile favorire un ingresso nella fase G0/G1 della coltura, per cui dopo lo splitting il primo ciclo risultera' parzialmente sincronizzato.
6 Deprivazione di siero. La deprivazione di siero puo' provocare in alcune linee cellulari una sincronizzazione (ma bisogna stare molto attenti a non indurre anche l'apoptosi). Burk ha infatti dimostrato che le cellule di Sirian hamster BHK21/C13 smettono di crescere se vengono trasferite in un medium che contiene 0.25% di siero, e possono essere mantenute in uno stato quiescente per 8 giorni o piu'. Ridando il siero non si ha nessuna sintesi di DNA per le successive 9 ore e un picco mitotico si puo' osservare tra le 23 e le 33 ore.
Sembra cosi' evidente che queste cellule si sono assestate in G1 e dopo la stimolazione hanno una lag phase di 9 ore prima di entrare nella fase esponenziale con un tempo di generazione di 10-12 ore.
Questo sistema va bene per cellule normali che vengono bloccate in G1. Le cellule tumorali invece, si bloccano spesso in varie fasi del ciclo, piu' per una mancanza di fattori nutritivi che per una vera e propria mancanza di fattori di crescita (dato che dovrebbero essere autocrine).
7 Deprivazione di isoleucina.
E' stato osservato che le cellule di ovaio di criceto (CHO) coltivate in terreno Ham's F10 e che sono entrate nella fase stazionaria, riprendono a dividersi se si cambia tipo di terreno. Questo e' dovuto al fatto che l'Ham's F10 e' un terreno molto povero di isoleucina (ne contiene circa il 5-10% rispetto agli altri terreni). Cellule come le CHO, L929, BHK21/C13, in crescita esponenziale, se trasferite in terreni poveri di leucina e con siero dializzato, si arrestano in G1. La sintesi di DNA crolla per parecchie ore anche se una duplicazione residua permane (dovuta anche alle cellule che gia' si trovavano nelle fasi S, G2 e M all'inizio del trattamento). Non sembra pero' esserci una crescita sbilanciata e le capacita' biosintetiche sono mantenute a livello sicuramente superiore che non nelle cellule entrate nella fase stazionaria. Nelle CHO l'aggiunta di isoleucina porta alla sintesi di DNA nella maggior parte delle cellule entro 4-5 ore, e entro 12 ore l'indice mitotico aumenta.
Il meccanismo d'azione della deprivazione di isoleucina non e' molto chiaro, ma non e' sicuramente dovuto ad una semplice mancanza di aminoacidi, dato che la mancanza di leucina, ad es., porta si' alla diminuzione della crescita, ma senza sincronizzazione. L'isoleucina sembra piuttosto avere un ruolo in qualche sottofase della G1 e sembra essere importante per l'azione dell'IGF.
Uno schema sperimentale puo' essere il seguente:
Raccogliere una sospensione di CHO in crescita esponenziale e lavarle in terrno Eagle's privo di isoleucina.
Seminare in piastre petri da 5 cm 7.5 x 105 cellule in Eagle's MEM senza isoleucina con 5% di FCS dializzato.
Lasciare una notte e quindi sostituire il terreno con medium fresco contenente isoleucina 2 x 10-6M e 10% di FCS non dializzato.
Questo metodo fornisce dei risultati comparabili alla selezione di cellule mitotiche con il 100% della popolazione e' in G1. E' un sistema piu' semplice della selezione mitotica ed e' piu' facilmente applicabile ad un maggior numero di cellule.
8 Sincronizzazione in fase S.
La sincronizzazione all'interfase G1/S viene ottenuta interferendo con la sintesi di uno o piu' desossirobonucleosidi trifosfati che sono richiesti per la sintesi del DNA, mentre le altre attivita' cellulari rimangono normali. Se si mantiene il blocco per almeno 16 ore circa il 70% delle cellule di una popolazione asincrona in fase esponenziale si viene a trovare all'inizio della fase S. In pratica non e' facile stabilire se le cellule non continuano comunque a sintetizzare DNA ma ad una velocita' estremamente lenta, cioe' se le cellule non siano comunque entrate nella fase S. Le cellule che erano gia' nella fase S al momento del trattamento vi rimarranno finche' il blocco non viene rimosso; a questo punto quel 70% di cellule che erano bloccate all'ingresso della fase S passeranno come una singola coorte attraverso la fase S e la G2, per poi dividersi. Queste cellule, molto utili per studiare gli eventi delle fasi S e G2, hanno pero' 2 svantaggi principali:
il primo e' che solo il 70% e' sincrono, l'altro 30% e' avanti di circa 6 ore.
- il secondo e' che il blocco della sintesi del DNA non e' detto che porti al blocco di altre attivita' che possono riguardare gia' la G1 o la G2. Inoltre con questo tipo di blocco aumenta la frequenza di rotture del DNA e di un inizio ripetuto di alcuni repliconi.
La crescita sbilanciata porta rapidamente alla morte cellulare se prolungata per piu' di una generazione. Una crescita sbilanciata si ha in ogni cellula che e' programmata a dividersi e quindi bloccata in una funzione, incluse le cellule mantenute in colcemide per piu' di poche ore. Inoltre il blocco della sintesi del DNA puo' avere effetti non apparenti finche' le cellule non vengono rilascite dal blocco e procedono vero la successiva G1.
La presenza di un 30% di cellule 'sfasate' puo' essere ovviato ad es. con un doppio blocco, cioe' l'agente che blocca la sintesi di DNA viene dato a cellule gia' bloccate in G1. La selezione iniziale puo' essere fatta con una selezione mitotica, oppure utilizzando cellule che siano appena state stimolate ad entrare nella fase di crescita, come linfociti stimolati con PHA o cellule comunque presincronizzate con uno dei trattamenti gia' descritti. Altri trattamenti sono l'uso di alte concentrazioni di timidina (ad es. trattando le cellule con timidina 3mM per 16 ore si ottiene il blocco del 70% di cellule al confine tra la G1 e la S. Dopo circa 8-10 ore tutte le cellule saranno diverse 'zone' della G1 e se le si tratta di nuovo con la timidina si bloccheranno di nuovo tutte le cellule che ora si trovano all'inizio della fase S). In pratica la sincronizzazione tra la fase G1 e la fase S viene ottenuta con molti reattivi, come timidina, aminopterina, ametopterina, 5-fluorodeossiuridina o idrossiurea. Quest'ultima e' molto usata, e il suo meccanismo d'azione si esplica inibendo una subunita' dell'enzima ribonucleotide reduttasi. Come tutte le sostanze sopracitate, agisce sul delicato equilibrio delle concentrazioni dei nucleotidi.
9 Procedure per indurre la sincronizzazione all'interfase G1/S.
1. Deprivazione di isoleucina e somministrazione di idrossiurea.
Piastrare 7.5 x 105 cellule in piastre da 5 cm in Eagle's MEM senza isoleucina e con 5% FCS dializzato.
Dopo 8 ore cambiare il terreno con Eagle's MEM con isoleucina e con 10% FCS non dializzato e idrossiurea 2 mM.
Dopo 16 ore cambiare il medium con medium completo senza idrossiurea. La maggior parte delle cellule saranno all'interfase tra G1 e S e cominceranno a replicare il DNA immediatamente.
2. Doppio blocco con timidina.
Piastrare 3 x 105 cellule HeLa in piastre da 5 cm in Eagle's Glasgow modificato contenente 10% FCS e timidina 3 mM.
Dopo 16 ore cambiare il medium con medium fresco senza timidina.
Dopo ulteriori 8 ore riaggiungere timidina fino alla concentrazione 3 mM. Dopo altre 16 ore ripetere il secondo passaggio, e a questo punto le cellule sono rilasciate dal blocco e entrano in fase S.
N.B. Questi tempi sono applicabili solo a cellule con un tempo di duplicazione di 24 ore. Non sono invece immediatamente trasponibili a cellule con tempi di generazione piu' brevi in cui la durata della fase S (tS) e' maggiore della meta' del tempo di duplicazione (T).
3. Deprivazione di siero e somministrazione di idrossiurea.
Un nostro protocollo utilizzato sempre sulle C108 prevede la deprivazione di siero per 24 ore e quindi il trattamento per 14 ore con idrossiurea 1.5 mM. La soluzione madre di idrossiurea e' una soluzione 25 X preparata in etanolo:PBS 1:1. Il rilascio dal blocco da idrossiurea viene ottenuto lavando le cellule 2 volte con PBS e rimettendole quindi in terreno completo.
9 Sincronizzazione in G2.
E' la fase in cui e' piu' difficile sincronizzare le cellule. Questo perche' le cellule sincronizzate o per selezione alla mitosi o per blocco all'interfaccia tra la fase G1 e la fase S arrivano in G2 gia' desincronizzate. La frazione massima ottenibile in G2 di CHO sincronizzate e' del 70% con il doppio blocco da timidina e del 40% con lo shaking mitotico. Un modo di sincronizzarle e' quello di avere dei mutanti termosensibili nella fase G2.
Tutti i metodi e le indicazioni riportate in questo capitolo sono state prese dal libro 'Cell culture for biochemists', di R.P.L. Adams, Elsevier, 1990. Questo libro fa parte della collana 'Laboratory Tecnhniques in Biochemistry and Molecular Biology', general editors R.H. Burdon and P.H. van Knippenberg. Non tutte le metodiche sono state riportate in questo testo per esteso.
|
|