|
| Appunti superiori |
|
|
| Appunti superiori |
|
| Visite: 2312 | Gradito: |
Leggi anche appunti:La cellulaLA CELLULA Le cellule furono osservate per la prima volta nel 1665 da Robert Il ciclo di krebsIL CICLO DI KREBS La maggior parte delle cellule eucariote e molti tipi DROSOPHILA MELANOGASTER (Drosophila melanogaster, fam: Drosophiline)DROSOPHILA MELANOGASTER (Drosophila melanogaster, fam: Drosophiline) Ø Obbiettivo: Osservare |
 |
 |
Fattore di trascrizione NF Y
Morfologia delle proteine NF Y
Il fattore di trascrizione "Nuclear Factor Y" (NF Y) è stato originariamente identificato in topo, come una proteina in grado di riconoscere il box Y nei promotori del Complesso Maggiore di Istocom patibilità di classe II (MHCII) Testa et al.,2005 . Questo fattore riconosce specificatamente l'elemento regolatorio CCAAT, ritrovato in entrambi gli orientamenti, in regioni prossimali e distali di promotori ed enhancer di molti geni, anche se nuovi studi di Chromatin Immuno Precipitation on chip (ChIP on- chip), hanno identificato un gran numero di loci genomici, il % del totale, distanti dal promotore che vengono legati in vivo da NF Y, presenti sia in introni che all'estremità ' o , ma l'associazione in vivo di NF Y ai promotori ed enhancer rimane essenziale per il funzionamento genico Guerra et al., 2007 .
Diversi studi genetici e biochimici condotti su mammiferi e piante hanno stabilito con certezza che gli attivatori CCAAT fanno parte di un complesso eterotrimerico NF Y, composto dalle tre subunità NF YA, NF YB, e NF YC, tutte richieste per il legame con la sequenza specifica consensu G/A, G/A, C, C, A, A, T, C/G, A/G, G/C. Testa et al., 2005 . In lievito è però presente una quarta subunità del complesso, detta Hap4, necessaria per l'attivazione trascrizionale soggetta a repressione da glucosio (in presenza di glucosio viene repressa, in assenza attivata) mentre le altre tre sono costitutive McNabb e Pinto, 2005 . Ogni subunità contiene una regione core, altamente conservata, suffic iente a creare sia il legame con la sequenza CCAAT che ad interagire con le altre subunità, le regioni fiancheggianti sono molto conservate, al contrario di quelle che includono i domini di attivazione. Lo studio dei domini conservati di NF YB e NF YC umani tramite cristallografia a raggi X (risoluzione 1,6 , ha permesso di capire come le proteine si relazionino tra loro, cioè tramite un "Histone folding motif" (HFM), tipico della struttura istonica che interagisce in un orientamento testa coda, suggerendo un'influenza reciproca diretta ma non specifica con il DNA, molto simile alle proteine istoniche istoni H2A H2B, rivelando poche differenze tra i loro motivi del core istonico formato da una struttura elica a - ansaL1 -elica a2- ansaL2- elica a3. La subunità NF-YA sembrerebbe avere un effetto di stabilizzazione del complesso, permettendo il legame sequenza specifico attraverso il dominio core di NF YA che, anche se lungo meno di 0 amminoacidi, risulta sufficiente per creare il legame con il DNA quando è complessato con il dimero NF- YC/NF YB. Il dominio core di NF YA si compone di due parti: un dominio N- terminale responsabile del legame NF YC/NF YB, e uno C-terminale, implicato nello specifico riconoscimento della sequenza CCAAT Mantovani et al.,1994 .
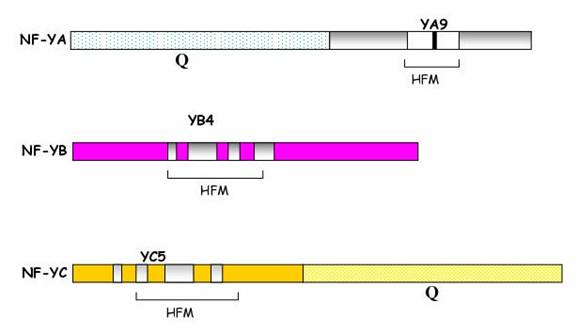
Fig . Rappresentazione schematica delle tre subunità costituenti il complesso eterotrimerico NF Y. HFM sta per Histone Fold Motif; YA9, YB4, YC5 sono le regioni core altamente conservate, e Q indica un dominio ricco in Q. Tratta da Guerra et. al., )
Molti studi hanno rivelato configurazioni comuni tra la coppia NF-YC/NF-YB con quella istonica H2A/H2B, dovute soprattutto alla presenza di elementi di struttura secondarie simili. Esaminando l'estremità carbossilica di entrambe le coppie proteiche, si vede come NF YB e NF YC abbiano le caratteristiche della rispettiva famiglia: NF YB, possiede, come H2B, un'elica aC, anche se risulta più corta, mentre in NF YC è presente un breve motivo ansa elica ansa nel HFM, simile alla breve elica a C ritrovata in H2A. Alcune differenze, però, spiccano tra gli amminoacidi che compongono le eliche, ad esempio la doppia catena intra arginina aspartato presente nell istone H3 (ma non in H2A e H2B), nella subunità NF YC è formata da residui assolutamente conservati di Arg 93 Ansa L2) e Asp 0 (elica a3), che non corrispondono in nessuna famiglia H2A, mentre per NF YB i residui Arg 8 ansa L2) e Asp 5 elica a 3 , anche essi assolutamente conservati, non sono presenti in H2B, dove vengono sostituita da una coppia di ripetizioni in arg/lys/glut. Un'altra particolarità di NF YC è nella posizione conservata del triptofano 85, alla fine dell elica a 2, racchiusa tra l'ansa L2 di NF YC e l'ansa L1 di NF YB, questa qualità è molto importante perché il residuo in questa posizione influisce con strutture superiori in questa regione. Vi sono differenze anche nell'assemblaggio del dimero: in quello istonico vi è da un lato, la terminazione C dell'elica a 3 di H2A e dall'altro l'ansa LC inizio elica a C di H2B, che attenua le interazioni con il resto del dimero, nel caso di NF YC, a C si ripiega indietro e sopra a 3 partecipando al grande core idrofobico formato dai suoi residui di a 2 e a 3 e dalla a 2 di NF YB, creando interazioni meno forti tra l ansa LC elica a C di NF YC e l'ansa LC elica a C di NF YB rispetto ad H2A/H2B, dove l'ansa LC inizio elica a C di H2B è vicino e interagisce fortemente con l'elica a C di H2A, grazie ad un glutammato conservato di H2A che imprime un effetto dipolo dall'elica a 2 di H2B Romier et al., 2003 .
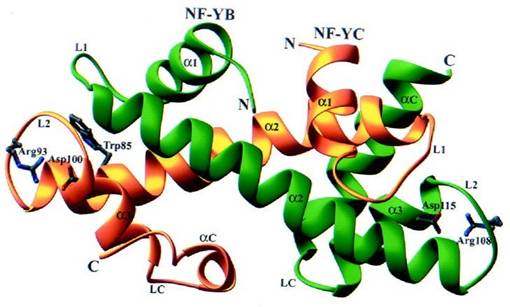
Fig : Struttura del dimero NF YB/NF YC
Vengono evidenziati i residui di Arginina e Asparagina comuni nei due peptidi e quello di Triptofano 5 presente nell'estremità a2 di NF C in grado di interagire con altre strutture superiori.
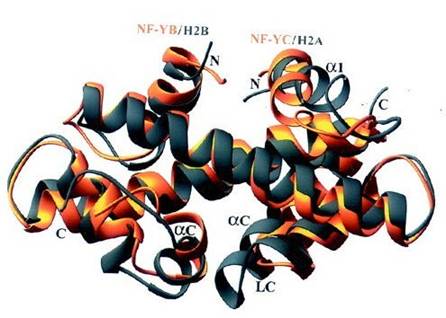
Fig. . Sovrapposizione delle strutture del dimero NF YB/NF YC con il dimero istonico omologo H2A/H2B.
2 Modalità di azione del complesso NF Y
Le regioni del core di NF YC e NF YB per potersi unire con l'ultima sub unità NF- YA devono soddisfare un prerequisito essenziale: devono associarsi in un eterodimero, rendendo così possibile la formazione della molecola trimerica. Una volta che il complesso trimerico si è formato, esso è in grado di legare il box CCAAT del DNA con un'alta specificità ed affinità, con una Kd tra le più alte dei fattori di trascrizione, compresa 10 0 e 1 M. Lo specifico riconoscimento delle basi sembra coinvolgere le interazioni con le anse maggiori e minori del DNA, e un saggio di permutazione circolare indica che, al disopra del legame, il DNA è incurvato di circa 60-80°, questa flessione potrebbe essere necessaria ad indicare l'orientamento esatto ai fattori di trascrizione Guerra et al., 2007 . L e regioni fiancheggianti al pentanucleotide CCAAT sono cruciali per un efficiente legame del complesso NF Y, anche se non è ancora noto come vengano riconosciute queste basi, cioè, se grazie alla sola distorsione del DNA o al riconoscimento delle regioni fiancheggianti o per entrambi i motivi. Molti studi biochimici hanno cercato di decifrare le diverse interazioni che intercorrono tra i fattori NF YA, NF YC, NF YB, e CCAAT box tra diverse specie, principalmente tra lieviti e mammiferi. Molti di questi studi si basano sull'analisi dei mutanti, dovuti a mutazioni puntiformi o delezioni, che possono destabilizzare il dimero o l'interazione dimero CCAAT principalmente in tre modi:
1. interferendo con l'assemblaggio del dimero NF YC/NF YB;
2. distruggendo l'effetto dipolo delle eliche a 1 e a 2 che permette il fissaggio dei gruppi fosfato;
3. abolendo le interazioni tra il dimero e il DNA, tramite impedimenti steric i o barriere elettrostatiche.
Due regioni del dominio core di NF YA sono state distinte come: regione N- terminale, detta NF YA1 di 234 257 residui, che riconosce il dimero NF YC/NF YB, e una regione C terminale, detta NF YA2 di 269-289 residui, responsabile del riconoscimento specifico del DNA. La regione N terminale di NF YA1 forma un a elica di cui hanno importanza funzionale i residui disposti solo da un lato dell'elica Romier et al.,2003 . Il sub dominio NF YA2 può essere suddiviso ulteriormente in una regione a elica N terminale (residui - ) e in una piccola regione C- terminale a spirale (residui 282- , come per NF YA1. In NF YB le mutazioni sull'elica a 2 (E90R e S97R), influenzano il legame con NF YA mentre in NF YC, entrambe le due eliche a 1 e a C si dimostrano importanti nella creazione del legame. Questi tre elementi di st ruttura secondaria sono presenti solo da un lato del dimero NF YC/NF YB, a formare un canale, dove avviene il legame con l'a elica N terminale di NF YA1, grazie anche a particolari residui amminoacidici di NF YA1, come Arg-245 e Arg- 249, che entrano in c ontatt o con altri residui presenti sulla superficie del dimero come Glu 90 e Ser 97. In questo modello la Ile 246 conservata di NF YA sarebbe compressa contro la Ile 117, diventando così esposta all ansa successiva dell elica aC di NF YC, è stato dimostrato che una mutazione che porta al cambiamento dell'ultima isoluecina in prolina, dissolve completamente il legame con NF YA evidenziando un estrema importanza della regione C terminale di NF YC nella formazionel del trimero Romier et al., 2003 . Diversi esperimenti di mutagenesi, inoltre, indicano che l'elica a 1 di NF YB influenza il riconoscimento del DNA da parte di NF YA2 e suggerisce che queste due regioni interagiscono direttamente, con l'elica a1 di NF YB probabilmente posizionata con l'elica NF YA 2 a in una corretta orientazione. Anche se l'orientamento dell'elica NF YA2 non è conosciuto perchè il linker che connette NFYA1 a NF YA2 può probabilmente occupare attraverso lo spazio sinistro tra il dimero NF-YC/NF-YB e il DNA o sorpassare il DNA. Il modello di interazione con il CCAAT box proposto (schematizzato in Fig. 5) offre due possibili localizzazioni per la sequenza di DNA: o l'elica a 1 di NF YB è posizionata esattamente al di sopra oppure su di un lato della sua ansa maggiore. Modellizazzioni della duplice interazione tra la parte dell'elica di NF YA2 con a1 elica di NF YB e il pentanucleotide CCAAT, sono difficile poiché i residui essenziali di NF YA2 possono essere coinvolti nello specifico riconoscimento delle basi, dello scheletro fosfato, oppure possono interagire con NF- YB (Romier et al., 2003
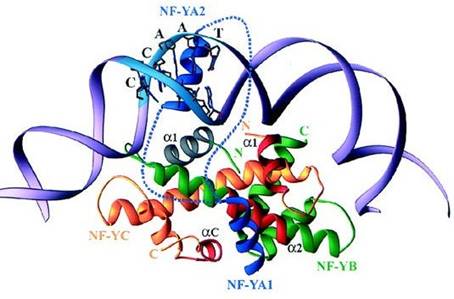
Fig. : Modello del complesso NF Y/CCAAT.
NF YC, NF YB e il DNA sono colorati rispettivamente in giallo, verde e viola , mentre NF YA è blu. Le due posizioni alternative pe r il link che connette i sub domini NF YA1 e NF YA2 sono in linea tratteggiata blu. Gli elementi della struttura secondaria della coppia istonica che sono coinvolti nel riconoscimento di NF YA1 e NF YA2 sono etichettati e colorati rispettivamente in rosso e grigio. Da Romier et. al., )
3 Tipologia di sequenza riconosciuta
L'importanza funzionale del motivo CCAAT box è emersa da numerosi studi riguardanti la caratterizzazione dei siti di inizio della trascrizione, condotti principalmente su promotori umani: questi hanno evidenziato che il CCAAT box o il suo revertito ATTGG è presente in più del % dei promotori, preferenzialmente localizzato nella regione 60, 100) rispetto al sito di inizio della trascrizione Testa et al., 2005 . Ulteriori analisi statistiche condotte su ottameri nucleotidi casuali, su
0 promotori umani, hanno inoltre confermato che la sequenza CCAAT è
seconda solo al Sp1 binding GC box in termini di abbondanza FitzGerald et al.,
. In generale, la posizione e l'orientamento dei nucleotidi fiancheggianti la regione centrale del penta nucleotide CCAAT sono estremamente conservati entro lo stesso gene tra le specie Guerra et al., 2007 . Negli organismi eucariotici, il pentanucleotide è presente a monte di geni sempre espressi, di geni inducibili Roy e Lee, ; Marziali et al., 997 , di geni tessuto specifici Berry et al., ; Ronchi et al., , di geni che controllano lo sviluppo e di geni che regolano il ciclo cellulare Frontini et alt,
4 Regolazione del fattore NF Y
Inizialmente era stato supposto che NF Y fosse un fattore trimerico costante, non inducibile, osservazioni più recenti hanno invece modificato questa convinzione. La capacità del fattore nucleare di legare il pentanucleotide CCAAT cambia, infatti, in stadi diversi di sviluppo o differenziamento cellule B, cellule muscolari in topo) e nella senescenza cellulare Yun et al., 3 . La regolazione di NF Y può avvenire a vari livelli, nei paragrafi successivi sono descritti alcuni esempi.
a REGOLAZIONE POST TRASCRIZIONALE
Le quantità degli mRNA relativi alle tre subunità rimangano sostanzialmente costanti, in diversi sistemi è stata rilevata una regolazione post trascrizionale che riguarda le subunità NF YA e NF YB. Si hanno infatti variazioni della disponibilità della subunità NF YA, in differenti tipi cellulari e/o condizioni di crescita Mantovani, ) ma non è chiaro se questo tipo di controllo si instaura sulla base della traduzione o della degradazione del peptide.
Sicuramente la NF YA costituisce la subunità regolatrice il complesso trimerico. La subunità NF YB invece, sebbene rimanga costante in quantità, potrebbe essere regolata attraverso mecanismi di fosforilazione e l'acetilazione.
b REGOLAZIONE DELLA LOCALIZZAZIONE NUCLEARE
Diversi fattori di trascrizione possiedono alcuni segnali caratteristici della loro localizzazione nucleari (NLS), in particolare questi segnali sono stati trovati nei quattro core istonici (H2A,H2B,H3,H4), nella porzione basica nell code N terminali e del HFM e sono mediati da un trasporto via importine. Studi biochimici indicano che NF YA e NF YB, sono presenti nel nucleo mentre NF YC è presente sia nel nucleo che nel citoplasma nei differenti stadi cellulari: nel citoplasma soprattutto durante la a fase G0 mentre viene rilocalizzato la durante la fase S Frontini et al., 2004 . Le subunità di NF Y sono importate nel nucleo grazie a distinti trasportatori della famiglia delle importine: l'importina ß media il trasporto di NF YA, mentre l'importina 13 media il trasporto del dimero NF YB e NF YC, poiché si associa solo quando i due HFM delle singole subunità sono uniti, diventando l'unico esempio conosciuto, per questo trasportatore, di substrato dipendente da lla formazione di un complesso Khale et al., 200 . L'importazione in forma di dimero avviene perché il singolo HFM contenuto nelle le subunità NF YB e NF YC non ha un NLS indipendente e di conseguenza in stato monomerico non sono né riconosciuti né importati da recettori di trasporto, ma in questa regione avviene anche il legame con la terza subunità NF YA, si crea così una competizione con l'importina13 ma il suo legame termina in presenza di un eccesso di NF Y A. Il particolare NLS di NF YA è nel dominio conservato della proteina al C terminale includendo la subunità che interagisce col DNA, consiste in più di 56 amminoacidi in cui i più importanti sono i residui di 11 arginine e 7 lisine la cui carica positiva è in grado di aumetare le probabilità di unirsi al recettore, mutazione di 6 dei 8 residui amminoacidici basici in questa regione causano la perdiata dell'accumolo della localizzazione nucleare.
Il trasporto mediato all'importina ß sembra essere circoscritto alla sola subunità
NF YA e non può essere esteso all'intero compleso trimerico. Gli HFM contenuti nel dimero non si comportano analogamente se sovraespressi: la sovraespressione di NF-YB non mostra una alterazione della localizzazione nucleare come avviene in NF YC.
Le proteine H2A simili come NF YC sono incapaci di interagire direttamente con il core istonici, mentre le proteine H2B simili come NF YB riescono a rapportarsi con H4 e H3, infatti NF YB a differenza di NF YC interagisce nel citoplasma con un'altra proteina nucleare e poi è trasportato nel nucleo. Allora, il trasporto a carico di NF YB nel nucleo potrebbe essere ottenuto via NLS nella coda N terminale del core istonico H3/H4. Inoltre, si è visto che il dimero NF YB/NF YC può efficientemente associarsi al DNA durante la formazione dei nucleosomi e questo complesso ha un'intrinseca affinità per H H4. Analisi di attivazione con l' HFM del dimero ha dimostrato che sono sufficienti ad attivare la trascrizione e altre prove biochimiche suggeriscono che NF YB e NF YC siano associati a differenti complessi ad alto peso molecolare in assenza di NF YA.
c "SPLICING" ALTERNATIVO
Le due subunità NF YA e NF YC possiedono, rispettivamente, due e tre isoforme principali generate da uno splicing alternativo, risultanti differentemente espresse in vari tipi cellulari. La forma breve di NF YA manca di 8 amminoacidi nel dominio attivo ricco in Q e presenta una piccola eterogenicità ai confini dei domini ricchi in Q e S/T. La forma breve più comune di NF YC è priva di 36 97 amminoacidi, codificanti per le eliche a1 e a3 del HFM Frontini et al., 2004 .
d REGOLAZIONE NELLE PIANTE
Un tipo di regolazione peculiare deve essere supposto per le piante che presentano famiglie multigeniche per tutte e tre le subunità del NF Y, a differenza degli altri eucarioti che presentano un gene per subunità.
In A. thaliana ad es. per tutte e tre le subunità esistono membri ubiquitari e altri strettamente tessuto specifici Gusmaroli et al., . I geni LEAFY COTYLEDON1 e LEAFY COTYLEDON1 LIKE, che appartengono alle NY B e sono espressi specificamente negli embrioni, saranno trattati nel capitolo 1.5.
|
| Appunti su: |
|