|
| Appunti superiori |
|
|
| Appunti superiori |
|
| Visite: 1634 | Gradito: |
Leggi anche appunti:Osservazione di una cipollaOsservazione di una cipolla OBIETTIVO: osservazione della struttura delle Sistemo endocrino ormoniSIST. ENDOCRINO. ORMONI: molecole organiche in una determinata regione dell' organismo LE CELLULE - Eterotrofi e autotrofi, Procarioti ed eucariotiLE CELLULE Eterotrofi e autotrofi L'energia che produsse le prime |
 |
 |
Effetti delle reazioni pro-ossidanti GGT dipendenti
Le specie reattive dell'ossigeno giocano un ruolo fondamentale nella modulazione di molte funzioni cellulari. In questo contesto la GGT e le reazioni proossidanti, conseguenti alla sua attività enzimatica, possono partecipare alla regolazione del metabolismo cellulare.
1 Azione modulatoria della GGT sui gruppi tiolici delle proteine
L'azione proossidante GSH/GGT dipendente può interferire con lo stato ossido-riduttivo dei gruppi tiolici delle proteine di membrana. Infatti queste sono il bersaglio primario delle molecole pro-ossidanti generate nell'ambiente extracellulare dall'azione della GGT. Il grado di ossidazione dei gruppi sulfidrilici delle proteine presenti sulla superficie cellulare è infatti aumentato in seguito alla stimolazione dell'attività di GGT mentre il processo è prevenuto dalla sua inibizione (Dominici et al., 2003). Il coinvolgimento di H2O2 in questo processo è indicato dal
fatto che l'ossidazione dei gruppi sulfidrilici delle proteine è significativamente prevenuta in presenza di catalasi.
L'ossidazione dei tioli proteici della superficie cellulare non sembra essere dovuta alla sola interazione di H2O2, prodotta dall'attività della GGT, con i gruppi sulfidrilici delle proteine (Radi, 1991; Quesada, 1996), ma è anche legata a processi di S-tiolazione che consistono nella formazione di un ponte disolfuro tra un gruppo
-SH proteico e un tiolo a basso peso molecolare (GSH, Cys, Omocys).
Molti dei gruppi sulfidrilici delle proteine si trovano nei siti attivi degli enzimi, la S-tiolazione ha quindi la duplice funzione di proteggere i sulfidrili delle proteine da un'ossidazione irreversibile e di modulare, in modo reversibile, la loro funzione (Coan et al., 1992; Schuppe-Koistinem et al., 1994; Sahaf et al., 2005). Infatti la "detiolazione", processo attraverso cui si ha la riduzione dei disolfuri misti, ne ripristina la funzione (Del Corso et al., 1993; Ravichandran et al., 1994, Cabiscol e Levine, 1996). Un numero sempre maggiore di molecole sembra essere modulato tramite S-tiolazione, sia proteine di membrana che proteine citosoliche (Sen e Parker, 2002; Sies e Parker, 2002) come ad esempio H-ras (Mallis et al., 2001), la chinasi p59 delle cellule T (Hehner et al., 2000), la fosfatasi PTP1B (Barrett et al.,
1999), c-jun (Klatt et al., 1999) NF-kB/p50 (Pineda-Molina et al., 2001) e la caspasi 3, implicata in processi apoptotici (Davis et al., 1997).
Anche la cisteinil-glicina originatasi dal catabolismo del GSH ad opera della GGT può concorrere alla formazione di ponti disolfuro con le proteine (Pompella et al., 2003; Corti et al., 2005). La S-cisteinilglicilazione così come la S-tiolazione può assumere due significati. In primo luogo, può essere interpretata come una difesa nei confronti dei danni ossidativi irreversibili (Coan et al., 1992; Thomas et al., 1995; Seres et al., 1996). In quest'ottica, essendo la GGT espressa ad alti livelli sia nei tumori che nelle metastasi (Tew et al., 1996) potrebbe contribuire alla resistenza delle cellule cancerogene nei confronti degli effetti citotossici dello stress ossidativo come nel caso di molti farmaci antitumorali ad attività proossidante (Daufeub et al.,
2002; Paolicchi et al., 2002b; Paolicchi et al., 2003). In secondo luogo la S-cisteinil- glicilazione può avere una funzione regolatoria. E' stato osservato che l'aumento della S-cisteinil-glicilazione comporta una diminuzione della S-glutatiolazione, ciò potrebbe essere considerato un meccanismo mediante il quale le cellule esprimenti attività di GGT, come le cellule tumorali (Tew et. al, 1996), riescono a modulare lo stato redox e la funzione di proteine importanti presenti nella matrice extracellulare e sulla superficie di altri tipi cellulari, quali per esempio le cellule del sistema immunitario o endoteliali (Corti et al., 2005).
2 Bersagli molecolari intracellulari della GGT: NF-kB AP-1 e TNFR1
NF-kB è un fattore di trascrizione ubiquitario implicato nella regolazione di un ampio numero di geni che controllano vari aspetti delle risposte immunitarie ed infiammatorie (Baeuerle e Henkel, 1994). Diverse sono le evidenze sperimentali a favore di una modulazione cellulo-specifica dell'attività di questo fattore di trascrizione in relazione all'attività di GGT. Utilizzando cellule di criceto V79-GGT che esprimono un transgene per GGT, è stato visto che la produzione di specie reattive dell'ossigeno GGT-dipendente promuoveva sia il legame di NF-kB al DNA sia l'attivazione del gene (Accaoui et al., 2000). Un altro studio su cellule di melanoma umano Me665/2/60 ha dimostrato che la stimolazione dell'attività dell'enzima o la sua inibizione determinavano rispettivamente la stimolazione o l'inibizione della traslocazione nucleare di NF-kB (Maellaro et al., 2000). Tuttavia in concomitanza a una aumentata attività dell'enzima è stato osservato un diminuito legame di NF-kB al DNA come se questo fosse un meccanismo di autoregolazione per prevenire un'eccessiva attivazione di questo gene in condizioni di persistente stress ossidativo (Dominici et al., 2003b).
Analogamente, l'attività di GGT sembra essere in grado di promuovere il legame di AP-1 al DNA; l'acivicina ma anche altri inibitori della attività di GGT sopprimono questo effetto a conferma del coinvolgimento dell'enzima (Paolicchi et al., 2002a).
Il tumor necrosis factor receptor I (TNFR1) è un recettore che presenta nel dominio extracellulare alcuni motivi ricchi in cisteine direttamente coinvolti nel legame con il tumor necrosis factor-α (TNF-α, Chen et al., 1995). Ciò suggerisce che la GGT modificando lo stato redox delle cisteine presenti sul TNFR-I possa influenzare l'affinità di legame del TNF-α al recettore e di conseguenza modulare le risposte cellulari a questa citochina (Dominici et al., 2004).
La possible modulazione del TNFR-I da parte dell'attività di GGT è stata studiata in due cloni della linea cellulari di melanoma Me665/2 caratterizzati da diversi livelli di attività enzimatica. In queste cellule sono state identificate cinque forme di TNFR-I distinte per il grado di ossidazione dei residui di cisteina. La forma più ossidata è costitutivamente presente nel clone cellulare 2/60, caratterizzato da un'elevata espressione di GGT, ed è possibile osservarla nel clone 2/21 in seguito all'induzione dell'espressione dell'enzima. La modulazione del recettore TNFR-I in seguito all'attività di GGT potrebbe modificare l'affinità di legame del TNFα, e quindi modulare la risposta cellulare (Dominici et al., 2004).
3 Effetti sulla proliferazione cellulare e apoptosi
L'apoptosi ha un importante ruolo biologico di regolazione nella maggior parte dei processi fisiologici come il differenziamento, l'omeostasi e la rimozione di cellule anormali in tutti i tessuti (Granville et al., 1998). Il controllo del meccanismo di morte cellulare programmata è modificato se non addirittura eliminato in cellule tumorali. E' nota da tempo l'interconnessione tra proliferazione cellulare, regolazione del ciclo e apoptosi, tanto che una alterazione del ciclo cellulare, peraltro inevitabilmente presente nelle cellule cancerose, può influenzare in maniera diretta la sensibilità della cellula agli stimoli apoptotici (Rudin e Thompson, 1997).
I dati raccolti nel corso di ricerche sulla linea cellulare di carcinoma ovarico umano A2780 hanno messo in evidenza come l'H2O2 e l'ossidazione dei tioli prodotti dall'attività di GGT durante il catabolismo del GSH svolgessero una azione antiproliferativa (Perego et al., 1997). Mentre, nella linea cellulare U937 di istiocitoma umano, Del Bello e collaboratori hanno osservato che la produzione di
specie ossidanti mediata dalla GGT costituiva uno stimolo antiapoptotico per le cellule e di conseguenza l'inibizione dell'enzima determini arresto della crescita e frammentazione del DNA caratteristica dell'apoptosi.
4 GGT nei tumori e resistenza a farmaci
La GGT è espressa in condizioni fisiologiche in numerosi organi e tessuti ed è stata trovata espressa ad alti livelli in lesioni maligne e premaligne nelle quali è considerata un fattore che conferisce un vantaggio per la moltiplicazione delle cellule neoplastiche. Inoltre l'attività di GGT sembra essere coinvolta nella resistenza delle cellule maligne ai farmaci alchilanti (Pompella et al., 2006).
Il vantaggio sulla crescita cellulare si pensa sia dovuto al fatto che la GGT, con la sua attività, favorisce la ricostituzione del GSH intracellulare (ciclo di Meister, Orlowski e Meister, 1970) e conseguentemente fornisce una protezione nei confronti del danno ossidativo. Inoltre, la GGT permette lo sfruttamento del GSH esxtracellulare come fonte di cisteina (ciclo di Meister, Orlowski e Meister, 1970), amminoacido essenziale e limitante la sintesi proteica. Come è stato già detto nei paragrafi 1-3, la GGT cellulare può influenzare il ciclo cellulare (proliferazione e apoptosi) e modulare l'attività di numerose proteine come recettori o fattori di trascrizione (TNFR1, NF-kB).
Il catabolismo del GSH extracellulare ad opera della GGT di membrana genera il dipeptide Cys-Gly e importanti osservazioni sono state fatte circa un ruolo di questo dipeptide nella resistenza ai farmaci antiblastici dei tumori che esprimono GGT. È stato dimostrato che la Cys-Gly formava un complesso con il cis- diammino-dicloro-platino (CDDP) riducendo la citotossicità del farmaco (Daubeuf et al., 2002, Paolicchi et al.2003).
5 Effetti della GGT sulle proteine LDL
E' stato discusso che in presenza di Fe2+ l'attività catalitica della GGT promuoveva la formazione di ROS i quali a loro volta potevano innescare il processo di perossidazione lipidica (Minotti, 1993). In studi successivi è stato verificato che le lipoproteine LDL erano ossidate in un sistema nel quale fossero presenti GGT, GSH, ADP-Fe3+ e GlyGly come accettori della reazione di transpeptidazione (Paolicchi et al., 1999). Gli esperimenti hanno mostrato che in miscele di reazione che includono
i complessi e GSH, quest'ultimo poteva ridurre il ferro ma la velocità di tale reazione aumentava significativamente in presenza di GGT e di GlyGly. Questo effetto è stato osservato in un intervallo esteso di attività di GGT (0-200 mU/ml) e di concentrazione di GSH (0-2 mM, Paolicchi et al. 1999).
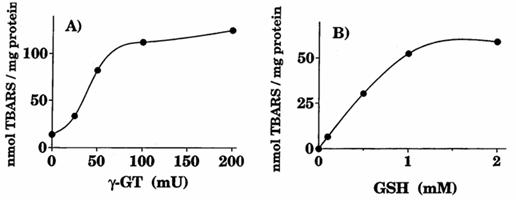
Fig 1.6 Ossidazione delle LDL dipendente dal GSH e dalla GGT
Incubazioni (1ml di volume finale) contenti LDL (0.1 mg prot/ml), Gly-Gly (20 mM) e ADP-Fe(III) (150 µM ADP-15 µM FeCl3) in PBS pH 7.4, 37°C. A)Le reazioni sono state iniziate mediante l'aggiunta di GSH (2mM), in più aumentando le concentrazioni di GGT. (B) GGT era mantenuta costante (50 mU/ml) e le reazioni sono state iniziate aggiungendo concentrazioni crescent di GSH. (Paolicchi et al., 1999)
Esperimenti aggiuntivi hanno indicato che l'ossidazione delle LDL dipendente dal GSH poteva essere promossa sia dalla GGT in presenza della trasferrina come sorgente di ioni ferro, sia da cellule esprimenti un elevato livello di attività di GGT sulla superficie, come ad esempio le cellule di epatoblastoma (HepG2) o le cellule monoblastoidi (U937) (Paolicchi et al., 1999). Le cellule in questo caso possono essere considerate come sorgente sia di GGT che di GSH, quest'ultimo infatti è normalmente rilasciato dalle cellule (Meister et al., 1995). Queste ultime due osservazioni sono una prima indicazione che l'ossidazione delle LDL mediata dal sistema GSH e GGT possa avvenire in condizioni fisiologiche.
|
| Appunti su: ossidazione residui cisteina mediata da h2o2, |
|