|
| Appunti scientifiche |
|
|
| Appunti scientifiche |
|
| Visite: 10777 | Gradito: |
Leggi anche appunti:Equazione di stato dei gas perfetti (Clapeyron)Equazione di stato dei gas perfetti (Clapeyron) Le tre leggi dei gas possono Equilibrio di solubilità, idrolisi e pHEquilibrio di solubilità, idrolisi e pH Il calcolo della solubilità di un Individuazione dei sali in un miscuglioINDIVIDUAZIONE DEI SALI IN UN MISCUGLIO OBIETTIVO: individuare |
 |
 |
Gli acidi nucleici sono l'acido desossiribonucleico (DNA) e l'acido ribonucleico (RNA). Prendono il loro nome dal fatto di essere presenti nel nucleo delle cellule.
La loro funzione è quella di contenere l'informazione genetica e renderla disponibile per guidare il metabolismo cellulare. Ogni organismo viene costruito e fatto funzionare a partire da un "progetto genetico" scritto nel suo DNA e reso operativo tramite il suo RNA.
Gli acidi nucleici presentano una struttura molecolare per molti versi analoga a quella delle proteine. Infatti, come le proteine, anche gli acidi nucleici sono costituiti da "collane" formate dalla successione ordinata di molecole più piccole. Ma mentre i mattoni che formano le proteine sono i 20 tipi di amminoacidi, gli acidi nucleici si formano a partire da 4 tipi di nucleotidi.
L'analogia tra la struttura molecolare delle proteine e quella degli acidi nucleici non è casuale. Le informazioni genetiche sono infatti "codificate" negli acidi nucleici come "successione dei nucleotidi". La cellula trasforma poi la successione dei nucleotidi in una ben precisa successione di amminoacidi. In altre parole, la particolare sequenza dei nucleotidi negli acidi nucleici rappresenta le informazioni genetiche necessarie per definire la struttura primaria di tutte le proteine di un organismo.
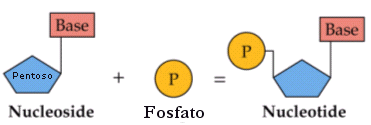
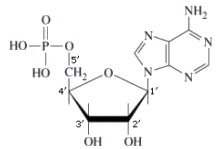
Un nucleotide è formato dall'unione di un pentoso che si lega ad una molecola di acido fosforico H3PO4 (gruppo fosfato) e ad una base azotata con legami di condensazione. In particolare si definisce nucleoside l'unione di un pentoso con una base azotata, mentre l'unione di un nucleoside con un gruppo fosfato genera un nucleotide (o nucleoside monofosfato).
Base + Pentoso = Nucleoside
Nucleoside + Fosfato = Nucleotide
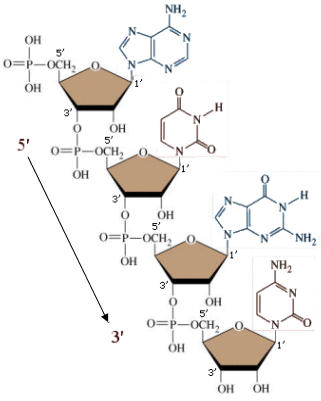
Il DNA è costituito da 4 tipi di nucleotidi diversi da quelli che formano l'RNA
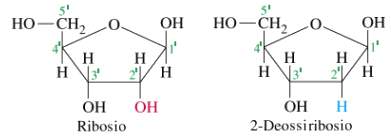
Nei 4 nucleotidi del DNA il pentoso è il desossiribosio (o deossiribosio), una molecola di ribosio che ha perso un atomo di ossigeno, mentre nell'RNA il pentoso è il Ribosio.
I 4 nucleotidi si differenziano tra di loro per le basi azotate (le basi azotate sono in questo analoghe ai gruppi R nelle proteine)
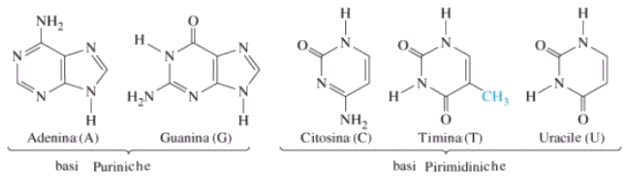
Le basi azotate del DNA sono:
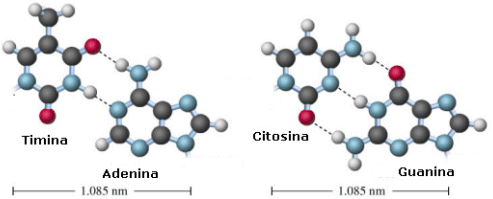
Adenina (A) Timina (T) Citosina (C) Guanina (G)
Le basi azotate dell'RNA sono:
Adenina (A) Uracile (U) Citosina (C) Guanina (G)
In altre parole i nucleotidi dell'RNA si differenziano da quelli del DNA solo per lo zucchero (ribosio al posto di deossiribosio) e per l'Uracile che sostituisce la Timina.
Adenina e Guanina sono basi puriniche
Citosina Timina ed Uracile sono basi pirimidiniche
Lo zucchero lega la base azotata in posizione 1' (uno-primo) ed il gruppo fosfato in posizione 5' (cinque-primo).
|
I 4 nucleotidi del DNA in forma libera (non legati nella catena del DNA) vengono indicati con le seguenti sigle: dAMP (deossiAdenosin Monofosfato) dGMP (deossiGuanosin Monofosfato) dCMP (deossiCitidin Monofosfato) dTMP (deossiTimidin Monofosfato) I 4 nucleotidi dell'RNA in forma libera vengono indicati con le seguenti sigle: AMP (Adenosin Monofosfato) GMP (Guanosin Monofosfato) CMP (Citidin Monofosfato) UMP (Uridin Monofosfato) I nucleotidi si legano tra loro in successione tramite legami fosfo-diesterei. L'ossidrile libero in posizione 3' (tre-primo) del pentoso viene utilizzato per legare il gruppo fosfato del nucleotide successivo. In questo modo si forma una catena in cui si succedono zucchero e fosfato, mentre le basi azotate rimangono sporgenti. La catena presenta quindi una precisa direzionalità, presentando ad un estremo il carbonio 5' (impegnato con un gruppo fosfato) ed all'altro estremo il carbonio 3' libero (non legato al gruppo fosfato). Le catene degli acidi nucleici vengono sempre montate in direzione 5' → 3'. |
|
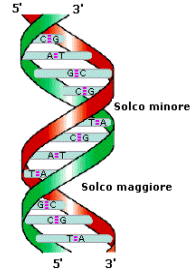
In natura il DNA si presenta con una struttura secondaria a doppia elica frutto dell'associazione di due singoli filamenti.
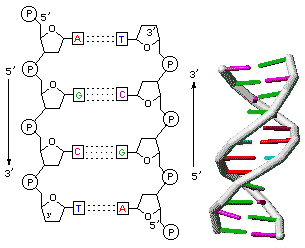
L'accoppiamento delle due catene si realizza attraverso la formazione di ponti a idrogeno tra le rispettive basi azotate, che vengono quindi a situarsi nella porzione centrale della struttura.
L'abbinamento fra i nucleotidi può realizzarsi soltanto tra basi cosiddette complementari, cioè fra adenina e timina, mediante due ponti a idrogeno (A=T), o tra citosina e guanina, mediante tre ponti a idrogeno (C≡G).
Le due catene, per fronteggiarsi, devono opporre una direzionalità inversa, essendo l'una diretta da 3' a 5', e l'altra, necessariamente, da 5' a 3', da cui è nato il termine di eliche antiparallele.
Le basi presentano una struttura planare e sono disposte in maniera parallela tra loro, come i gradini di una scala a chiocciola.
La complementarietà dei due filamenti ha una conseguenza fondamentale. Infatti, qualora sia stabilita l'esatta successione di basi di un filamento, si può ricavare, seguendo semplicemente le leggi della complementarietà, la serie di nucleotidi del filamento corrispondente.
In questa conformazione ciascuna coppia di basi dista dalle contigue 0,34 nm. Un giro completo dell'elica, o passo dell'elica, misura 3,4 nm, per cui sono presenti 10 coppie di basi per un giro completo. Il diametro dell'elica è di 2nm. L'avvitamento è destrorso, ossia, immaginando di guardare lungo l'asse, i due filamenti si avvolgono in senso orario.
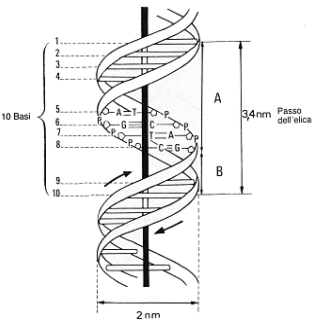
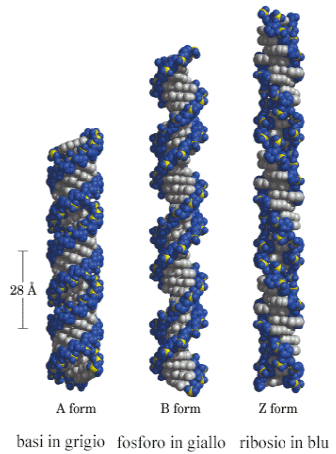
La struttura del DNA in natura ed in condizioni normali viene detta di tipo B con le basi perpendicolari all'asse dell'elica. La struttura B è la più frequente ed è quella a cui ci si riferisce in generale. Rappresenta il DNA in una soluzione acquosa e forma al suo esterno due solchi di diverse dimensioni chiamati solco maggiore e solco minore.
In condizioni particolari esistono altre due strutture dette A e Z.
La struttura A non esiste in vivo e corrisponde alla molecola disidratata ed è più compatta. Contiene 11 basi per giro, presenta un diametro di 2,4 nm ed un passo di 2,6 nm.
La struttura Z è un elica sinistrorsa con lo scheletro che presenta un andamento ondulato (Z = Zig-zag).
Contiene 12 basi per giro, presenta un diametro di 1,8 nm ed un passo di 3,7 nm. Questa struttura si può formare in brevi tratti costituiti da una alternanza delle basi G e C ripetute più volte. Si pensa che possa avere delle funzioni strutturali e di informazione in alcuni processi cellulari e inoltre una sua eccessiva presenza sembra essere correlata alla comparsa di malattie autoimmuni.
Come abbiamo già detto il DNA contiene il "progetto costruttivo" ed i "programmi di gestione e manutenzione" di un organismo, codificati nella sua struttura primaria (sequenza basi). Le sue funzioni, strettamente correlate alla sua natura di deposito di informazioni, sono:
Rendere disponibili le informazioni genetiche per la costruzione ed il corretto funzionamento dell'organismo. Tale funzione viene mediata da molecole di RNA che copiano le informazioni genetiche (trascrizione) e le trasformano (traduzione) in proteine (sintesi proteica). Le diverse sequenze di nucleotidi, caratteristiche del DNA di un organismo, definiscono infatti quali amminoacidi debbano succedersi nella costituzione delle sue proteine. Tra le proteine sintetizzate vi sono naturalmente anche gli enzimi, prodotti per controllare le reazioni di cui la cellula necessita. In questo modo il DNA controlla, attraverso la sintesi degli enzimi, tutto il metabolismo cellulare
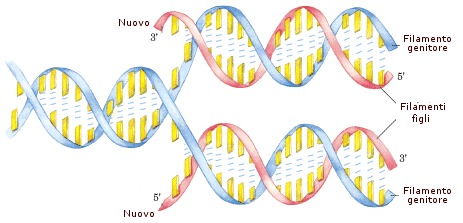
Generare copie delle informazioni genetiche (duplicazione o replicazione del DNA) in modo da rendere disponibile l'intero progetto genetico per le nuove cellule che si formano. Ogni essere vivente pluricellulare nasce infatti a partire da una sola cellula (zigote), la quale ha il compito di moltiplicarsi fino a formare i miliardi di cellule di cui è composto l'organismo adulto. Al momento di ogni divisione cellulare (mitosi) devono dunque essere già predisposte due copie del materiale genetico (DNA) in modo che ogni nuova cellula possieda per intero tutte le informazioni per funzionare correttamente.
Tutte le funzioni svolte dal DNA sono rese possibili grazie alla complementarietà delle basi azotate.
Duplicazione del DNA
Nel processo di duplicazione (o replicazione) del DNA, il doppio filamento viene attaccato e tagliato in un punto dall'enzima topoisomerasi. La doppia elica viene successivamente aperta come una cerniera-lampo dall'enzima elicasi che, rompendo i ponti ad idrogeno che tengono unite le basi azotate complementari, separa i due filamenti e forma una struttura ad Y detta forcella di replicazione. La forcella di replicazione viene stabilizzata dalle proteine SSB. Un altro enzima (DNA-polimerasi) provvede poi ad agganciare su ognuno dei due filamenti esposti (filamenti-genitori o parentali) dei nucleotidi complementari. In questo modo su ciascun filamento viene ricostruito il filamento mancante (filamento-figlio) e si generano due copie della doppia elica originaria. Poiché in ciascuna di queste due copie sopravvive metà della molecola originaria, tale processo è anche noto come sintesi semiconservativa.
Per le caratteristiche intrinseche al legame stesso, la costruzione dei nuovi filamenti avviene solo in direzione 5'→3'. Ed avendo il DNA due filamenti antiparalleli, la sintesi dei due nuovi filamenti avviene necessariamente in direzioni opposte.
Un filamento, detto filamento veloce (leading strand) viene sintetizzato in modo continuo, poiché la DNA-Polimerasi avanza nella stessa direzione dell'elicasi.
L'altro filamento, detto filamento lento (lagging strand) viene sintetizzato in modo discontinuo, con la formazione di segmenti di DNA detti frammenti di Okazaki, che vengono successivamente saldati dall'enzima DNA-ligasi.
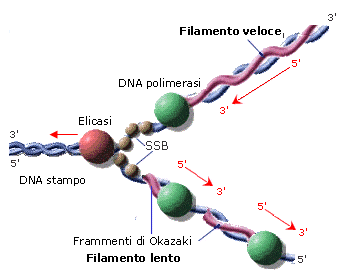
La DNA-polimerasi necessita di un innesco, cioè di una breve sequenza di RNA (RNA primer) da cui partire per sintetizzare il nuovo filamento, dal momento che è in grado solo di aggiungere nucleotidi a una catena preesistente, che fornisca una estremità 3'-ossidrilica libera. Inoltre la DNA-polimerasi possiede un secondo sito attivo in grado di controllare che l'attività di appaiamento delle basi sia avvenuta correttamente e di correggere eventuali errori. Nel caso vengano individuati errori nel processo di duplicazione la DNA-polimerasi torna indietro a correggerli. Questa funzione di correzione è definita attività 3'-5' esonucleasica (proofreading activity = correzione di bozze).
La duplicazione del DNA avviene contemporaneamente in più punti della molecola, con le forcelle di replicazione che si allontanano in direzioni opposte formando bolle di replicazione, destinate ad estendersi ed a fondersi.

Mentre la bolle di replicazione crescono i filamenti di DNA non ancora aperti tendono a manifestare una tensione torsionale aumentando il numero di spire per unità di lunghezza (si pensi a cosa accade ad una treccia se tiriamo le estremità dei fili divaricandoli). Le topoisomerasi di classe I risolvono il problema della tensione causato dall'avanzamento dell'elicasi tagliando periodicamente uno dei due filamenti e permettendo all'elica di girare, per svolgere gli avvolgimenti in eccesso. Quando il DNA si è rilassato, la topoisomerasi riconnette il filamento rotto, ripristinando il DNA a doppia elica.
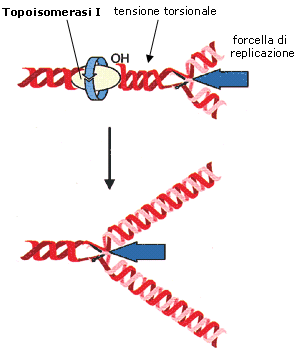
I frammenti di Okazaki vengono successivamente saldati dalla DNA ligasi, mentre i primers di RNA vengono rimossi e sostituiti da analoghi filamenti di DNA da un enzima della famiglia delle DNA-polimerasi.
Come abbiamo già visto l'RNA è costituito da una catena di nucleotidi in cui lo zucchero è il ribosio e la Timina è sostituita dall'Uracile. La complementarietà delle basi è in questo caso Adenina-Uracile e Citosina-Guanina.
Mentre il DNA è una molecola di grandi dimensioni e notevolmente stabile, gli RNA sono più piccoli, hanno vita limitata e inoltre sono caratterizzati da una struttura a singolo filamento.
Le molecole di RNA possono ripiegarsi su se stesse in modo tale da permettere la formazione di legami ad idrogeno tra basi complementari appartenenti a tratti diversi e distanti della medesima catena con formazione di zone a struttura secondaria elicoidale.
Piccole molecole di RNA sono quindi in grado di acquisire una struttura tridimensionale regolare, spesso responsabile della loro funzione specifica, mentre RNA più grandi presentano zone a struttura tridimensionale definita, congiunte fra loro da parti non strutturate.
Le molecole di RNA vengono sintetizzate utilizzando un filamento di DNA come stampo sul quale vengono appaiati i singoli nucleotidi complementari dell'RNA (trascrizione).
Spesso la forma funzionale di un RNA è più corta rispetto al trascritto primario. Il passaggio dalla forma primaria inattiva a quella finale funzionale avviene ad opera di enzimi specifici che rimuovono alcune zone della molecola (introni) e saldano quelle rimanenti (esoni) attraverso un processo di taglia-incolla detto splicing.
L'RNA permette al DNA di trasformare le informazioni in esso contenute in proteine. Esistono 3 tipi di RNA, tutti coinvolti nella sintesi proteica.
Come abbiamo già detto il DNA contiene le informazioni necessarie per posizionare nella giusta successione gli amminoacidi di una proteina. Il tratto di DNA che "codifica" per una particolare proteina si definisce "gene". Le informazioni "genetiche" sono codificate nella struttura primaria del DNA, sono cioè scritte nella successione delle sue basi azotate.
Nel descrivere la sintesi proteica si utilizza una metafora linguistica, in cui esistono due linguaggi: quello del DNA con un alfabeto di 4 lettere (le basi azotate) e quello delle proteine con un alfabeto di 20 lettere (gli amminoacidi).
Le informazioni devono pertanto essere "tradotte" da un linguaggio ad un altro. Ovviamente per effettuare la traduzione e "decifrare" un "messaggio codificato" è necessario possedere il "codice" che fornisce la corrispondenza tra i simboli dei due linguaggi.
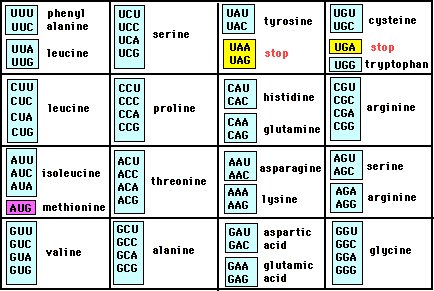
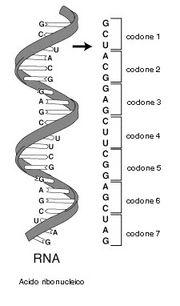
Il "codice genetico" definisce dunque il modo in cui la successione delle basi azotate del DNA deve essere tradotta nella corretta successione di amminoacidi di una proteina. Ovviamente non vi può essere una corrispondenza biunivoca tra basi azotate ed amminoacidi (4 contro 20). Si è scoperto che il "vocabolario" del DNA è formato da 64 "parole", formate dalla combinazione delle quattro "lettere" A T C G (basi azotate) prese a gruppi di tre (43 = 64).
Ciascun amminoacido viene dunque "codificato" da una particolare tripletta di basi o codone. I codoni sono le triplette già trascritte nella molecola dell'RNA messaggero e quindi con l'Uracile al posto della Timina.
Esistono più triplette che codificano per il medesimo amminoacido (ridondanza del codice genetico), ma ciascuna tripletta non può, ovviamente, codificare per amminoacidi diversi.
Esistono anche triplette che non codificano per alcun amminoacido, dette triplette "not-sense", che la cellula utilizza come "segni di interpunzione" durante la traduzione dell'informazione per segnalare la fine (stop) della sintesi della proteina.
Il processo di sintesi proteica si articola in due fasi: trascrizione e traduzione dell'informazione genetica. Nella fase di trascrizione l'informazione viene trasferita dal DNA all'RNA, mentre nella fase di traduzione l'informazione passa dall'RNA alle proteine
In questa fase l'informazione genetica viene copiata (trascritta) dal DNA su di una molecola di RNA. Il processo di trascrizione avviene grazie all'enzima RNA-polimerasi. Nelle cellule eucarioti ci sono tre diverse molecole di RNA-polimerasi, che occupano diversi siti. Ciascuno di questi enzimi è responsabile della trascrizione di una differente classe di geni.
L'RNA-polimerasi I, che risiede nel nucleolo, è responsabile della trascrizione dei geni per la produzione di tutto l'RNA ribosomiale (o rRNA). Questo è l'enzima con la più elevata attività di sintesi.
L'RNA-polimerasi II, localizzata nel nucleoplasma (la parte di nucleo che esclude il nucleolo), responsabile della sintesi del precursore dell'RNA messaggero (mRNA).
l'RNA-polimerasi III, l'enzima con l'attività minore, anch'essa presente nel nucleoplasma, che sintetizza l'RNA di trasporto (tRNA).
Nella fase di inizio l'RNA-polimerasi si lega alla doppia catena del DNA, aprendola in corrispondenza di una particolare sequenza, chiamata promotore. Il promotore è una speciale sequenza di nucleotidi che non verrà trascritta, situata sul DNA all'inizio del gene.
Successivamente l'RNA-polimerasi scorre lungo il DNA rompendo i ponti Idrogeno tra le basi azotate complementari ed aprendo la doppia elica come una cerniera. In questo modo una delle due catene viene esposta alla copiatura e fa da stampo per la sintesi di una molecola di RNA messaggero ad essa complementare. Mentre l'RNA-polimerasi scorre sul filamento-stampo del DNA vengono agganciati ad esso dei ribonucleotidi complementari. Quando, durante la trascrizione, nel DNA si incontreranno particolari sequenze di basi alla fine del gene (terminatore) si avrà il termine della trascrizione. Il filamento di RNA messaggero si stacca ed il DNA si richiude e si riavvolge
Poiché i due filamenti si legano tramite appaiamento delle basi azotate complementari, questi sono tra loro antiparalleli. La direzione di lettura del DNA è 3'→5' mentre quella di trascrizione è 5'→3'.
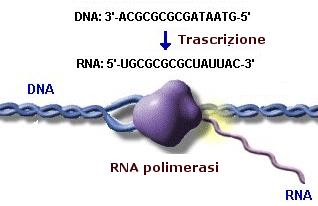
Il prodotto della trascrizione è denominato trascritto primario e consiste probabilmente in un filamento di RNA che si estende dal promotore al terminatore. Non si ha dimostrazione di ciò perché esso è molto instabile e quindi difficile da isolare.
La fase cruciale della produzione delle diverse forme di RNA è la maturazione a partire dai precursori.
I complessi trascritti primari degli rRNA e tRNA di procarioti ed eucarioti vengono modificati in forme mature più semplici. Gli mRNA dei procarioti non subiscono quasi mai modificazioni, mentre l'assemblaggio dell'mRNA degli eucarioti è piuttosto complesso.
Negli eucarioti la trascrizione genera dei precursori nucleari degli mRNA (trascritti primari) caratterizzati dalla presenza di zone non codificanti o introni (intron = intragenic region). Tali precursori vengono in seguito convertiti negli mRNA maturi attraverso un processo (splicing) che prevede la rimozione degli introni e il ricongiungimento delle parti codificanti o esoni (exon = expressed region). Lo splicing avviene grazie a un apparato enzimatico complesso (spliceosoma) in grado di riconoscere sequenze specifiche presenti nelle zone di giunzione esone-introne, di rimuovere gli introni e di ricongiungere correttamente tra loro i vari esoni.
Una volta maturati, gli mRNA, come le subunità ribosomiche e i tRNA, passano nel citoplasma per svolgere la loro funzione nella sintesi proteica.
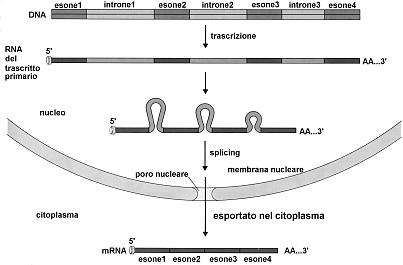
Le tappe della maturazione avvengano tramite la modificazione delle due estremità della molecola (Capping e Poliadenilazione) e la rimozione degli introni (Splicing)
Il 5' capping (rivestimento in 5') consiste nell'apposizione di un cappuccio all'estremità 5' (5' cap) del trascritto primario, costituito da un GTP (Guanosin Trifosfato) modificato (la Guanina presenta un gruppo metile in posizione 7). All'estremità 5' dell'RNA messaggero viene asportato il gruppo fosfato ed il GTP modificato si salda ad essa con un inusuale ponte trifosfato 5'-5'.
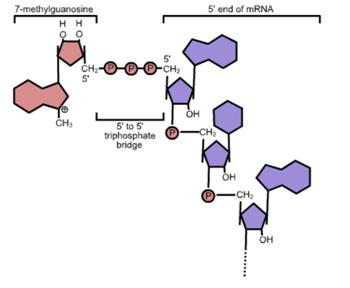
Il cappuccio in 5' ha la funzione di
promuovere il successivo processo di traduzione costituendo il sito di attacco per il ribosoma
prevenire la degradazione dell'RNA ad opera delle esonucleasi dato che il rivestimento in 5' assomiglia ad una estremità 3'
La poliadenilazione è una fase della maturazione del trascritto primario che consiste nell'aggiunta di una sequenza di 150-250 AMP all'estremità 3'. La coda di poliadenina sembra essere essenziale sia nel garantire stabilità all'RNA messaggero che nei processi di uscita dal nucleo e di traduzione.
Lo splicing avviene grazie all'azione di un gruppo di riboproteine chiamate snRNP (small nuclear RiboNucleoProteins o Snurps). Una snRNP è costituita da una molecola di RNA associata a proteine.
Le molecole di RNA che formano le snRNP sono note come snRNA (small nuclear RNA = piccolo RNA nucleare). Essendo molto ricche in Uracile sono state siglate U1, U2, U4, U5 e U6. Le proteine che entrano nella formazione delle snRNP sono note come Ribonucleoproteine Sm (Sm sta per Smith).
Le snRNP riconoscono e agganciano sequenze specifiche presenti nelle zone di giunzione esone-introne e ne catalizzano i processi di taglio e ricongiunzione . I siti enzimatici sono localizzati sulle molecole di snRNA anziché sulle proteine, come avviene di norma, e quindi le snRNP sono definito come ribozimi.
Un ribozima (crasi dei due termini 'acido ribonucleico' + 'enzima'), o enzima a RNA o RNA catalitico, è una molecola di RNA in grado di catalizzare una reazione chimica.
Secondo alcuni autori i ribozimi sarebbero una sorta di "fossili molecolari", un'eredità di un ancestrale mondo a RNA, dove l'Acido Ribonucleico era la molecola in grado sia di portare l'informazione che di effettuare la catalisi. L'ipotesi del mondo a RNA (W.Gilbert - 1986) è una teoria che propone la presenza di forme di vita basate esclusivamente sull'RNA prima della formazione degli attuali organismi viventi basati soprattutto sul DNA.
Gli introni presentano le seguenti caratteristiche comuni, che permettono alle snRNP di riconoscerli:
una sequenza GU in corrispondenza del sito di taglio 5' (5' splice site)
una sequenza AG in corrispondenza del sito di taglio 3' (3' splice site)
un sito di ramificazione A (branch site) in prossimità dell'estremità 3'contenente una Adenina
un segmento di polipirimidine (Py tract) ricco in pirimidine (Citosina ed Uracile) tra il sito di ramificazione e l'estremità 3'
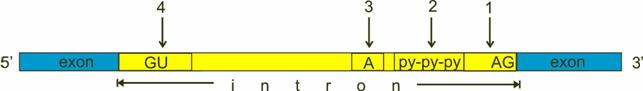
Lo splicing inizia con la snRNP-U1 che riconosce e aggancia il sito di taglio 5', mentre la snRNP-U2 riconosce e aggancia il sito di ramificazione.
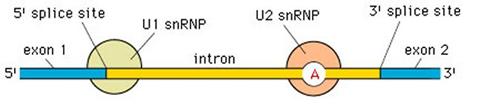
Il successivo aggancio delle altre snRNP (U4/U6 e U5) costringe l'introne a formare un cappio in modo che le sue estremità di taglio si avvicinino. L'apparato enzimatico costituitosi, formato dal complesso di tutte le snRNP è detto spliceosoma.
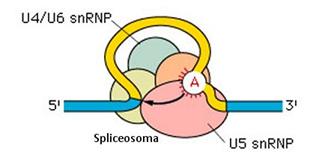
Lo spliceosoma taglia l'estremità 5' e connette l'estremo libero dell'introne all'Adenina del sito di ramificazione, andando a formare un cappio chiuso noto come 'lariat'.
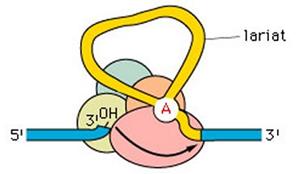
Successivamente verrà tagliata anche l'estremità 3'. Lo spliceosoma ricongiungerà le due estremità esoniche e si distaccherà asportando l'introne in forma di lariat.
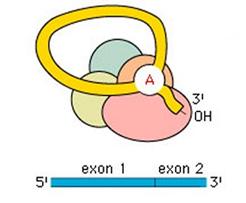
L'RNA messaggero (mRNA) rappresenta la classe di RNA più eterogenea; infatti è costituita da filamenti contenenti tanti codoni quanti sono gli amminoacidi delle proteine da loro codificate.
RNA messaggeri codificanti per piccole proteine sono costituiti da alcune centinaia di nucleotidi, quelli codificanti per proteine grandi ne comprendono varie migliaia. Ogni mRNA è caratterizzato dal codone d'inizio (spesso AUG, specifico per l'amminoacido metionina). I tre codoni UAA, UGA e UAG rappresentano invece il segnale di terminazione della sintesi della catena polipeptidica. La precisione nell'andamento lineare dei ribonucleotidi in gruppi di tre, non solo determina il corretto allineamento degli amminoacidi in una proteina, ma anche un esatto punto di inizio e di conclusione della sua sintesi.
L'RNA di trasporto (tRNA) trasferisce ai ribosomi i vari amminoacidi che, uniti tra loro con legame peptidico, formano le proteine. Molti trascritti primari che originano dai geni per i tRNA sono discretamente più lunghi rispetto alle piccole molecole mature che si riversano nel citoplasma e che contengono molte basi modificate. Come tutte le macromolecole trasportate dal nucleo al citoplasma, anche i tRNA maturi vengono trasportati attraverso i pori nucleari, probabilmente associati a proteine specifiche che ne facilitano il passaggio. Una volta giunti nel citoplasma, i tRNA maturi si presentano come molecole piccole, costituite da 75-80 nucleotidi che si appaiano tra loro in zone specifiche con ponti idrogeno tra basi complementari, interrotte da tratti a singolo filamento. Tale situazione determina una particolare conformazione a "trifoglio", caratteristica per tutti i tRNA. Nella cellula, tuttavia, questa molecola ha una complessa organizzazione a forma di L rovesciata e contorta a spirale, poiché le due anse laterali del trifoglio si avvicinano tra loro formando l'angolo fra i bracci della L. L'estremità 3' del filamento polinucleotidico di tutti i tRNA sopravanza quella 5' di tre nucleotidi uguali (C-C-A). Tale sequenza rappresenta il sito accettore dell'amminoacido. L'amminoacido viene saldato in coda del tRNA dall'enzima amminoacil-sintetasi, con formazione du un amminoacil-tRNA. Si distinguono circa venti tRNA, ciascuno specifico per un determinato amminoacido.
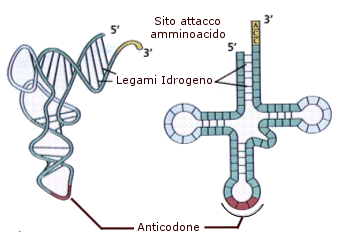
La parte più caratteristica della molecola del tRNA è l'ansa terminale, detta anticodone poiché porta tre basi complementari ai codoni degli mRNA.
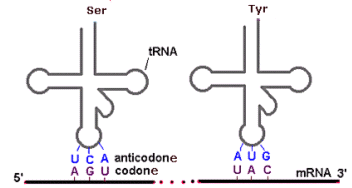
Gli RNA ribosomiali (rRNA) costituiscono una famiglia di molecole che, assemblate insieme a più di 50 diverse proteine, formano i ribosomi. I ribosomi sono gli organuli citoplasmatici che utilizzano le informazioni genetiche dell'RNA messaggero e gli amminoacidi portati dagli RNA di trasporto per assemblare le proteine. Sono costituiti da due subunità classificate in termini di Svedberg (S), una misura del coefficiente di sedimentazione di particelle in sospensione sottoposte a centrifugazione (gli organuli cellulari vengono separati tramite centrifugazione in base alla loro diversa densità). La lunghezza delle molecole di rRNA, la qualità delle proteine costituenti ciascuna subunità e di conseguenza la grandezza di queste ultime varia tra procarioti ed eucarioti.
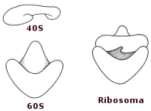
In base ai loro coefficienti di sedimentazione, i ribosomi sono stati suddivisi in due classi:
- I ribosomi 70 S sono caratteristici dei procarioti e sono formati da una subunità 30 S e da una 50 S.
- I ribosomi 80 S sono caratteristici degli eucarioti e sono formati da una subunità 40 S e da una 60 S
Negli eucarioti i geni che codificano per gli rRNA sono localizzati nel nucleolo, che si evidenzia come un corpicciolo sferico situato nel nucleo. Tale conformazione è dovuta all'intensa attività trascrizionale che si attua al livello di questi geni e dal quasi contemporaneo assemblaggio degli RNA alle proteine ribosomiali.
ll ribosoma, presentando molecole di RNA in grado di catalizzare la formazione del legame peptidico, è considerato un ribozima.
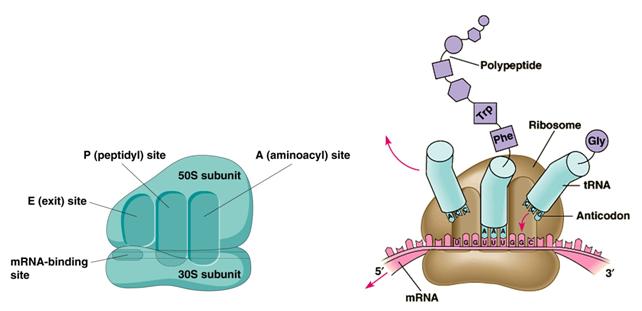
La subunità minore funge da sostegno e da sito di ingresso dell'mRNA da tradurre.
Nei ribosomi procarioti la subunità minore (30S) presenta un sito di aggancio (RBS - Ribosome Binding Site) costituito da una breve sequenza di basi (AGGAGG - sequenza di Shine-Dalgarno) complementare alla sequenza che nel filamento di mRNA precede il codone di inizio (AUG) della traduzione.
Nei ribosomi eucarioti la subunità minore (40S) scansiona il filamento di mRNA, a partire dalla sua estremità 5', finchè non riconosce una sequenza d'inizio (sequenza di Kozak) contenente il codone di inizio (AUG) della traduzione.
La subunità maggiore è la principale macchina catalizzatrice del complesso. Quando sono unite le due subunità possiedono tre siti d'attacco per i tRNA:
il sito A che rappresenta il sito di aggancio (Acceptor) per gli Amminoacil-tRNA in entrata
il sito P che è normalmente occupato dal Peptidil-tRNA, cioè dalla molecola di tRNA che porta il filamento proteico in via di allungamento
Il sito E (Exit) che rappresenta il sito di uscita del tRNA ormai scarico.
Ciascun tRNA che entra nel ribosoma va ad occupare in successione tutti e tre i siti.
Nella fase di traduzione, l'informazione genetica, contenuta nell'RNA messaggero come sequenza di codoni (triplette di basi), viene letta dai ribosomi in direzione 5' → 3' e trasformata nella corrispondente sequenza di amminoacidi, in direzione N-terminale → C-terminale, grazie al codice genetico.
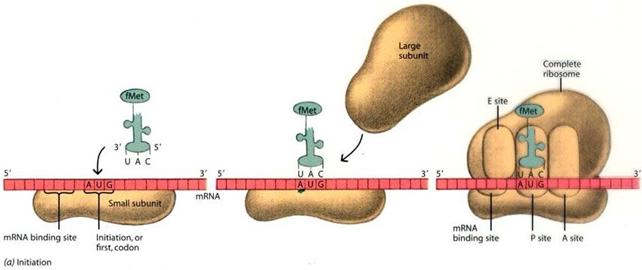
Nella fase di inizio (initiation) la subunità minore del ribosoma aggancia l'estremità 5' dell'mRNA e si posiziona sul codone d'inizio (AUG) della traduzione. Successivamente si posiziona la subunità maggiore.
Il primo amminoacil-tRNA ad essere aggiunto, detto tRNA iniziatore, è invariabilmente quello legato all'amminoacido metionina, complementare al codone d'inizio, il quale si posiziona nel sito P. La metionina viene poi rimossa alla fine della traduzione da una proteasi specifica.
Nella successiva fase di allungamento (elongation) della catena proteica Il secondo amminoacil-tRNA si posiziona nel sito A. In questo modo il suo amminoacido risulta adiacente a quello portato dal primo amminoacil-tRNA
Il sito catalitico della subunità maggiore (peptidiltransferasi) salda con legame peptidico i due amminoacidi, rompendo contemporaneamente il legame tra l'amminoacil-tRNA che occupa il sito P ed il suo amminocido.
Il ribosoma scivola sul filamento di mRNA in direzione 3', scalando di un codone. In questo modo i due tRNA che occupavano i siti P ed A, vanno ad occupare rispettivamente i siti E e P, mentre il sito A si libera.
Mentre un nuovo amminoacil-tRNA entra ad occupare il sito A, il tRNA scarico presente nel sito E si sgancia ed esce dal ribosoma, lasciando il suo amminoacido sulla catena proteica in via di formazione agganciata al tRNA che occupa il sito P, detto peptidil-tRNA.
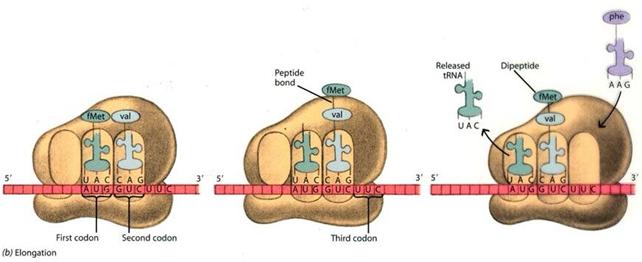
La traduzione procede con il medesimo meccanismo, con il ribosoma che avanza di un codone per volta ed un tRNA che entra carico del suo amminoacido ed uno che esce scarico del suo amminoacido. Un ribosoma umano aggiunge circa 2 amminoacidi al secondo.
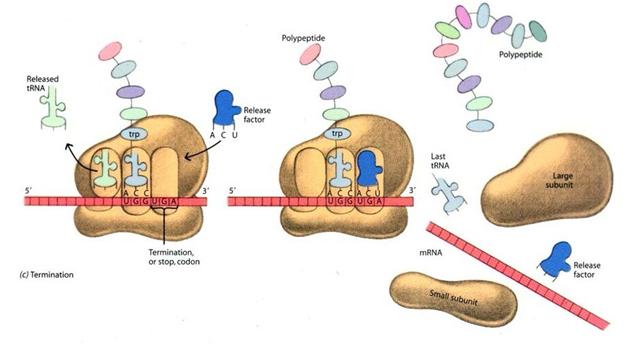
Il processo termina (termination) quando il ribosoma trova un codone di stop, in corrispondenza del quale si lega un fattore di rilascio. Il filamento proteico si stacca e viene liberato nel citoplasma dove assume la sua conformazione nativa.
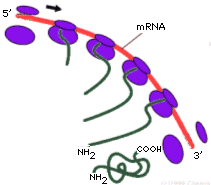 Un filamento di mRNA può essere letto e tradotto più volte in modo da
ottenere più copie della medesima proteina. La traduzione può essere fatta
contemporaneamente da più ribosomi che si infilano sul medesimo filamento come
le perle di una collana. La struttura che ne deriva prende il nome di poliribosoma (o polisoma). Ciascun ribosoma appartenente ad un polisoma si trova
ovviamente in una fase diversa della sintesi della medesima proteina
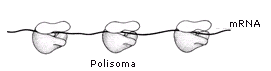
Un filamento di mRNA può essere letto e tradotto più volte in modo da
ottenere più copie della medesima proteina. La traduzione può essere fatta
contemporaneamente da più ribosomi che si infilano sul medesimo filamento come
le perle di una collana. La struttura che ne deriva prende il nome di poliribosoma (o polisoma). Ciascun ribosoma appartenente ad un polisoma si trova
ovviamente in una fase diversa della sintesi della medesima proteina
ATP e NAD sono due nucleotidi modificati che la cellula utilizza per il suo metabolismo energetico.
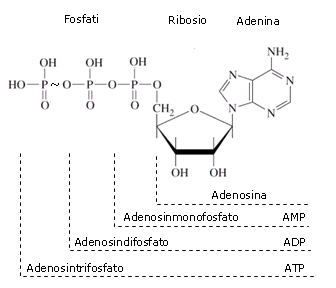
L'ATP (Adenosin Trifosfato) viene utilizzato dalle cellule come contenitore e trasportatore di energia. L'ATP deriva dall'AMP (Adenosin Monofosfato = Adenina-Ribosio-Fosfato) per aggiunta di due molecole di acido fosforico.
Nel primo passaggio si forma ADP (Adenosin Difosfato = Adenina-Ribosio-Fosfato-Fosfato) secondo la seguente reazione di condensazione che avviene tra il gruppo fosfato dell'AMP e la molecola di acido fosforico
AMP + H3PO4 → ADP + H2O
Spesso nei composti organici il gruppo fosfato viene indicato, in modo abbreviato, con il simbolo Pi (Fosforo inorganico). La reazione di formazione dell'ADP può quindi essere scritta in forma sintetica
AMP + Pi → ADP
L'ATP si forma dalla reazione di condensazione di una molecola di ADP con un'altra molecola di acido fosforico. La reazione è fortemente endoergonica e richiede circa 7,3 kcal/mol (30,5 kJ/mol)
ADP + Pi + 7,3 kcal → ATP
L'ATP risulta in tal modo una molecola altamente energetica. L'energia è fissata nel legame tra il secondo ed il terzo gruppo fosfato, legame che viene rappresentato con un tratto circonflesso
Adenina-Ribosio-Pi-Pi~Pi
La cellula utilizza l'energia estratta dai suoi combustibili (carboidrati e lipidi) per sintetizzare ATP. In questo modo l'energia non viene dissipata sotto forma di calore, ma viene fissata come energia di legame in molecole, come l'ATP, rapidamente utilizzabili per fornire energia. Ad esempio, la combustione di una molecola di glucosio permette alla cellula di caricare 36 ATP
C6H12O6 + 6O2 → 6CO2 + 6H2O + Energia (36ATP)
Quando la cellula ha bisogno di energia può ottenerla in modo rapido semplicemente idrolizzando ATP
ATP → ADP + Pi + 7,3 kcal
Durante il metabolismo cellulare questa molecola oscilla dunque continuamente tra la forma carica (ATP) e la forma scarica (ADP)
Il NAD (Nicotinammide AdeninDinucleotide) è una ossidoreduttasi, un coenzima che catalizza reazioni di ossidoriduzione. Il NAD è quindi in grado di trasferire elettroni (ed Idrogeno) da una molecola che si ossida ad una molecola che si riduce. Ovviamente quando il NAD acquista elettroni da una molecola A che si ossida il NAD passa in una forma ridotta (NADH o NAD ridotto), mentre quando cede elettroni ad una molecola B che si riduce passa in una forma ossidata (NAD+ o NAD ossidato)
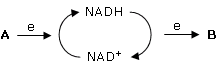
La reazione di riduzione del NAD richiede 2 elettroni, uno ione H+ e 52,4 kcal/mol (220 kj/mol)
NAD+ + 2e + H+ + 52,4 kcal → NADH
In modo analogo a quanto accade per l'ATP, anche il NAD oscilla quindi tra una forma scarica (NAD+) ed una forma carica (NADH).
Altre importanti ossidoreduttasi analoghe al NAD sono il FAD (Flavin Adenin Dinucleotide) e il NADP (NAD fosfato).
Analogamente al NAD esistono entrambi nella forma ossidata e ridotta
FAD ossidato (FAD) e FAD ridotto (FADH2)
NADP ossidato (NADP+) e NADP ridotto (NADPH)
Il FAD è in grado di assorbire un po' meno energia del NAD
FAD + 2e + 2H+ + 46,0 kcal → FADH2
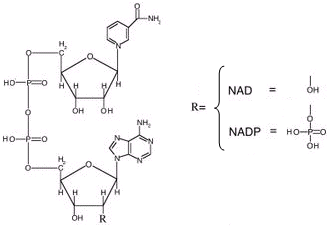
NAD e FAD hanno come precursori alcune vitamine del gruppo B. Il NAD deriva dalla vitamina B3 (o niacina o vitamina PP = Prevenzione Pellagra), il FAD dalla vitamina B2.
|
| Appunti su: poliribosoma, dna gruppo fosforico, desossiribosio, struttura secondaria dna, gruupo fosfato, |
|
| Appunti Geografia |  |
| Tesine Biologia |  |
| Lezioni Ingegneria tecnico |  |