|
| Appunti scientifiche |
|
|
| Appunti scientifiche |
|
| Visite: 9160 | Gradito: |
Leggi anche appunti:Bilanciamento con numeri di ossidazione frazionariBilanciamento con numeri di ossidazione frazionari I numeri di ossidazione Numero di ossidazione (nox) o stato di ossidazione (stox)Numero di ossidazione (nox) o stato di ossidazione (stox) Si definisce numero Uso dell'equazione cinetica integrataUso dell'equazione cinetica integrata Se si possiedono una serie di misure |
 |
 |
La cellula eucariote animale è formata da una membrana plasmatica esterna che racchiude il citoplasma ed il nucleo.
Il citoplasma è costituito da una frazione liquida, il citosol, e dagli organuli cellulari in essa sospesi.
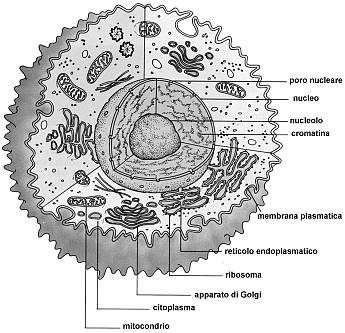
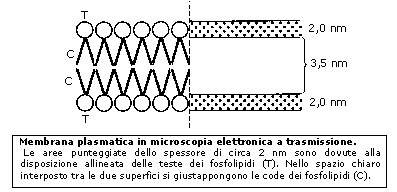
La membrana plasmatica (o plasmalemma) racchiude il contenuto della cellula e costituisce una barriera fra l'ambiente intracellulare (ambiente interno) e quello extracellulare (ambiente esterno). È costituita da un doppio strato continuo di fosfolipidi, dello spessore di 8-10 nm (un nanometro corrisponde a un miliardesimo di metro = 10-9 m), attraversata in parte o completamente da numerose proteine. È inoltre presente una piccola percentuale di glucidi, in forma di glicoproteine e glicolipidi, e di molecole di colesterolo che la stabilizzano, I fosfolipidi sono lipidi formati da una 'testa' di glicerolo legato ad un gruppo fosfato e a due 'code' di acidi grassi. Come conseguenza della presenza di acqua sia all'interno che all'esterno della cellula, i fosfolipidi rivolgono le teste polari verso le due superfici della membrana, mentre le code apolari sono rivolte verso la parte interna del doppio strato
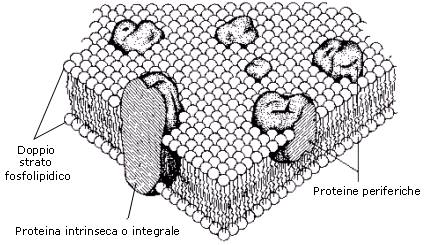
In condizioni fisiologiche, sia le molecole lipidiche sia quelle proteiche in esse immerse sono in grado di muoversi liberamente (modello a mosaico fluido). Le caratteristiche di fluidità consentono alla membrana di manifestare proprietà autosigillanti attraverso le quali può autoripararsi, ossia richiudersi rapidamente ed automaticamente qualora in essa si produca un foro, automaticamente. La capacità autosigillarsi permette inoltre alle membrane di fondersi e scindersi, senza che si abbiano perdite di materiale cellulare, nel corso di processi fondamentali per la vita e l'attività delle cellule (divisione cellulare, esocitosi, endocitosi).
La membrana rappresenta l'interfaccia della cellula con l'ambiente esterno, attraverso la quale avvengono e vengono regolati tutti gli scambi di materiali ed informazioni. Essa ha dunque funzione di trasporto, di interazione ed integrazione con l'ambiente e di locomozione.
La membrana plasmatica è una barriera selettivamente permeabile. Il doppio strato fosfolipidico permette infatti il libero passaggio solo dell'acqua, di gas come O2 e CO2 e di piccole molecole liposolubili, mentre risulta impermeabile per ioni e molecole idrosolubili.
Il trasporto dei materiali per i quali la membrana non è permeabile viene effettuato dalla cellula utilizzando particolari proteine trasportatrici transmembraniche (carriers) e/o attraverso fenomeni di motilità della membrana (esocitosi ed endocitosi).
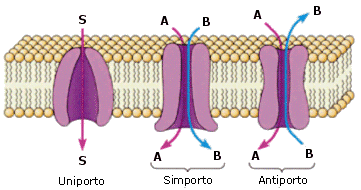
Le proteine trasportatrici sono proteine in struttura terziaria in grado di riconoscere in modo specifico la sostanza chimica da trasportare (substrato).. Esistono tre modalità di trasporto:
Uniporto: una sola molecola in un unico verso
Simporto: due molecole in un stesso verso
Antiporto: due molecole in versi opposti, una in ingresso e l'altra in uscita
Le proteine trasportatrici si dividono in pompe e canali
1) Le pompe presiedono al trasporto attivo di molecole. Nel trasporto attivo il passaggio di una molecola avviene con consumo di energia (idrolisi ATP), anche contro il gradiente di concentrazione.
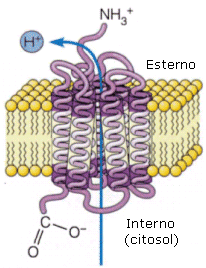
Batteriorodopsina
La batteriorodopsina è una pompa protonica (protone = ione H+) che alcuni batteri utilizzano per catturare l'energia luminosa. È una proteina transmembranica costituita da 7 alfa-eliche in associazione con una molecola di retinale
2) I canali presiedono al trasporto passivo di molecole. Nel trasporto passivo il passaggio di una molecola avviene per diffusione senza consumo di energia, secondo il gradiente di concentrazione, La diffusione è il fenomeno fisico per il quale un fluido (gas o liquido), come conseguenza del moto disordinato delle molecole che lo costituiscono, tende a "sparpagliarsi" occupando tutto lo spazio disponibile. La conseguenza, a livello macroscopico, è la tendenza spontanea di un fluido a muoversi (diffondere) da zone a maggior concentrazione verso zone a minor concentrazione. Si dice appunto che tale movimento avviene "secondo il gradiente di concentrazione". Il fenomeno si giustifica in termini di probabilità.
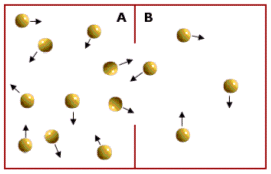
Le particelle si muovono in tutte le direzioni e quindi in ogni istante alcune passeranno dalla regione A alla regione B e viceversa. Ma poiché vi sono più particelle in A che in B vi sarà una maggior probabilità che si muovano da A verso B, piuttosto che da B verso A. Dunque, finché le particelle non saranno distribuite omogeneamente vi saranno più particelle che passano da A verso B che particelle che passano da B verso A. Ciò genera un movimento netto verso B (diffusione) che porta ad eliminare la distribuzione spaziale non omogenea.
Ovviamente i canali possono funzionare solo se tra l'interno e l'esterno della cellula esiste una differenza di concentrazione.
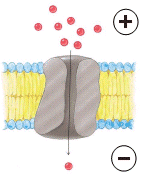
La cellula è in grado di aprire o chiudere i suoi canali a seconda delle necessità.
Avvalendosi di questi meccanismi di trasporto, la cellula può mantenere la concentrazione interna delle sostanze chimiche su valori diversi da quelli che caratterizzano l'ambiente esterno.
Come conseguenza delle caratteristiche di semipermeabilità della sua membrana, la cellula è soggetta a fenomeni osmotici. L'osmosi è un fenomeno che si manifesta quando una membrana selettivamente permeabile (semipermeabile) separa due soluzioni a diversa concentrazione. La membrana è tale per cui solo le molecole dell'acqua possono liberamente diffondere, mentre la diffusione del soluto è impedita. In queste condizioni l'acqua tende a diffondere dalla soluzione più diluita (dove è presente in quantità maggiore) alla soluzione più concentrata (dove è presente in quantità minore). Il flusso dell'acqua verso la soluzione più concentrata produce una vera e propria pressione (pressione = forza/superficie = forza che agisce sull'unità di superficie), detta pressione osmotica. Il valore della pressione osmotica è direttamente proporzionale alla differenza di concentrazione tra le due soluzioni. Nel caso le due soluzioni poste a contatto siano contenute in recipienti aperti il flusso d'acqua dall'una all'altra produrrà un dislivello che potrà essere utilizzato come misura della pressione osmotica (ricordiamo che 10 metri d'acqua equivalgono alla pressione di 1 atmosfera).
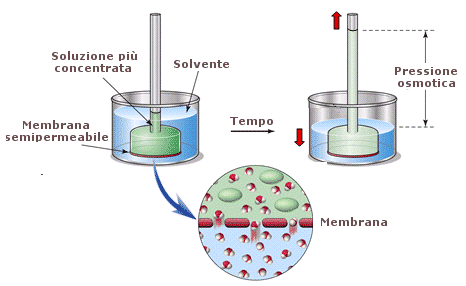
I fenomeni osmotici possono creare problemi alle cellule animali. Poste in un ambiente ipertonico (tono = concentrazione; ipertonico = a concentrazione maggiore) possono facilmente disidratarsi. Poste in un ambiente ipotonico (a concentrazione minore) possono assorbire acqua fino alla lisi cellulare. Ne sono un esempio i nostri globuli rossi che sono immersi in un mezzo isotonico (plasma). Le cellule vegetali che, esternamente alla loro membrana plasmatica, sono rivestite di una robusta parete di cellulosa, sono invece in grado di sfruttare i fenomeni osmotici. Le cellule vegetali mantengono infatti il loro citosol leggermente ipertonico rispetto all'ambiente circostante. In questo modo richiamano acqua per osmosi e risultano soggette ad una leggera pressione, detta turgore cellulare, che le mantiene gonfie. Il turgore cellulare è fondamentale per mantenere distesi i tessuti vegetali verdi, come le foglie, che non presentano strutture di sostegno efficaci.
3) Alcune cellule eucarioti sono in grado di importare ed esportare considerevoli quantità di materiali sfruttando la motilità e la fluidità della loro membrana, la quale è in grado di "deformarsi" producendo introflessioni ed estroflessioni. Tali deformazioni della membrana sono note come movimenti ameboidi (l'ameba è un organismo unicellulare in cui tale caratteristica è particolarmente sviluppata).
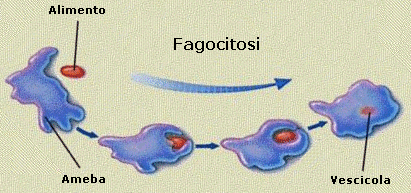
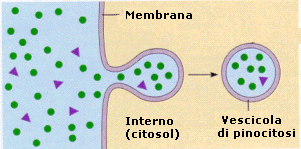
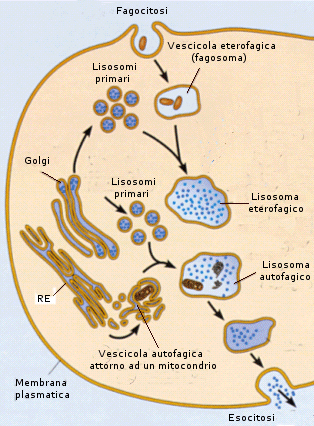
Il processo di importazione è detto endocitosi ed avviene grazie alla capacità della cellula di avvolgere il materiale da introdurre con una piccola porzione di membrana che si stacca come una goccia (vescicola di endocitosi o endosoma) all'interno del citosol. Si parla di endocitosi mediata da recettori quando la sostanza che deve essere inglobate nella cellula, viene riconosciuta e legata da specifici recettori situati sulla superficie della membrana. Si viene a formare in questo modo un complesso recettore-ligando che funge da attivatore del processo di endocitosi. Se il materiale da introdurre è solido si parla di fagocitosi, se è liquido di pinocitosi.
La fagocitosi avviene grazie alla crescita di due estroflessioni, dette pseudopodi (pseudopodio = falso piede), attorno al materiale da inglobare. I pseudopodi crescono fino ad avvolgere il materiale e a fondersi alle sue spalle inglobandolo in una vescicola di fagocitosi. In questo modo, ad esempio, i macrofagi, un particolare tipo di globuli bianchi del nostro organismo inglobano e distruggono i batteri ed eliminano cellule morte e strutture estranee.
La pinocitosi si produce grazie alla formazione di minuscole introflessioni della membrana all'interno delle quali viene risucchiato il liquido da introdurre. La membrana successivamente si richiude liberando nel citosol la vescicola di pinocitosi
Il processo di esportazione è detto esocitosi ed avviene grazie alla capacità della cellula di avvolgere il materiale da espellere con una piccola porzione di membrana (vescicola di esocitosi) che si stacca dai sistemi interni di membrana (apparato del Golgi) e si va a fondere con la membrana plasmatica. In tal modo il contenuto delle vescicole di esocitosi viene versato all'esterno.
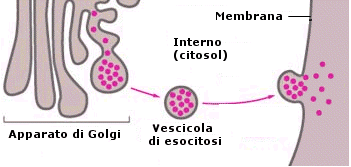
La cellula utilizza l'esocitosi sia per eliminare le sostanze di rifiuto (escrezione), ma anche per effettuare la secrezione di sostanze utili, come segnali chimici (neurotrasmettitori, ormoni etc) o sostanze aggressive per attaccare altre cellule (enzimi digestivi, tossine etc)
La cellula può utilizzare la motilità di membrana anche per la locomozione, per muoversi cioè su di un supporto. In questo caso il movimento ameboide avviene con la crescita di uno pseudopodio nella direzione del movimento. Una volta raggiunta la sua massima estensione lo pseudopodio fa presa sul supporto e la cellula viene richiamata in avanti (un po' come fanno le lumache, per intenderci).

Ovviamente la locomozione è una caratteristica che si trova prevalentemente in organismi unicellulari. Le cellule appartenenti ad un organismo pluricellulare sono strettamente addossate l'una all'altra e non si muovono. Fanno comunque eccezione alcune cellule del sistema immunitario (macrofagi e granulociti).
I movimenti ameboidi non sono l'unica possibilità di locomozione per la cellula. Alcune cellule presentano infatti sulla loro membrana delle estroflessioni citoplasmatiche filiformi (ciglia e flagelli) che possono essere utilizzate per la locomozione. Di tali strutture parleremo più avanti.
La membrana rappresenta anche il mezzo con cui la cellula si 'fa riconoscere' dalle altre cellule, tramite molecole dette marcatori, riconosce segnali chimici, tramite molecole dette recettori, è "sensibile" agli stimoli tramite la sua eccitabilità e si collega ad altre cellule negli organismi pluricellulari tramite giunzioni proteiche.
1) I marcatori di membrana sono in genere proteine legate a carboidrati (glicoproteine), che funzionano come una sorta di 'carta d'identità' in base alla quale la cellula viene riconosciuta come facente parte del sé (self), ossia dell'organismo stesso, e non viene attaccata dal sistema immunitario, oppure come estranea (not self) e come tale, da distruggere. L'insieme delle molecole che caratterizzano i diversi tipi di cellule e di tessuti dell'organismo viene chiamato complesso maggiore di istocompatibilità (MHC). Esso è responsabile del fatto che i tessuti trapiantati agiscono da antigeni e vengono attaccati dall'organismo ricevente (fenomeno del rigetto).
2) I recettori di membrana sono in genere anch'essi costituiti da molecole proteiche in grado di riconoscere e legarsi in modo specifico e selettivo a particolari sostanze chimiche che operano come segnali, principalmente nervosi (acetilcolina, adrenalina, serotonina etc) ed ormonali. Una volta legati, il complesso recettore-segnale innesca un meccanismo di amplificazione dovuto all'azione di un enzima legato al recettore che provoca una cascata di reazioni all'interno della cellula.
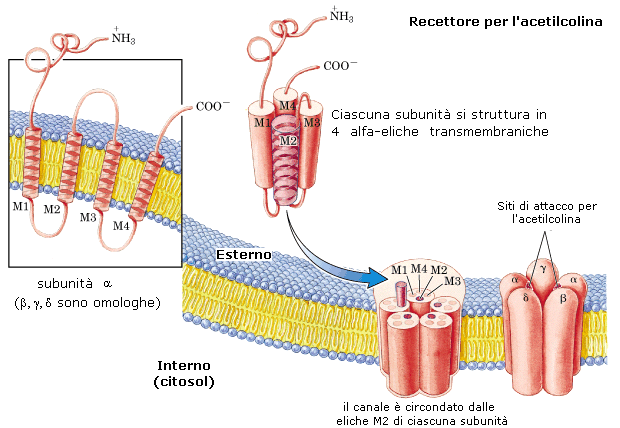
3) Eccitabilità. In condizioni normali la membrana cellulare risulta polarizzata, con un eccesso di ioni positivi sulla superficie esterna ed un eccesso di ioni negativi sulla superficie interna. La differenza di potenziale tra le due facce è di circa -70 milliVolt, (il segno negativo è convenzionale) ed è chiamato potenziale di riposo. Questa stato di polarizzazione è alla base della capacità della cellula di percepire stimoli esterni e di reagire ad essi. Questa "sensibilità" della cellula viene definita eccitabilità. La membrana si eccita quando, come conseguenza di uno stimolo, inverte il suo potenziale (depolarizzazione) da -70 a circa +30 mV (potenziale di azione). L'eccitabilità di membrana è alla base del funzionamento delle cellule nervose e muscolari. Il potenziale di riposo viene generato dall'azione congiunta della pompa Sodio-Potassio e dei relativi canali.
La pompa Na+/K+ é una proteina intrinseca che lavora associata ad una ATPasi che, idrolizzando ATP, libera l'energia necessaria per il trasporto (trasporto attivo). In presenza di ATP, la pompa cambia conformazione e si lega internamente con 3 ioni Na+ ed esternamente con 2 ioni K+. Ad ogni idrolisi di una molecola di ATP, 3 ioni sodio vengono pompati verso l'esterno della cellula, mentre 2 ioni potassio verso l'interno. La membrana cellulare è praticamente impermeabile allo ione sodio (i canali del Sodio sono chiusi), che rimane quindi per lo più segregato nello spazio extra-cellulare, mentre è permeabile allo ione potassio (i canali del potassio sono aperti), che è quindi libero di ridiffondere all'esterno. Questa distribuzione ineguale di cariche elettriche fa si che l'interno della cellula sia carico negativamente rispetto all'esterno caricato positivamente.
4) In un organismo pluricellulare le cellule si collegano l'una all'altra mediante giunzioni intercellulari. Nelle piante superiori le cellule sono connesse mediante 'ponti' di citoplasma denominati plasmodesmi.
Nella maggior parte degli animali, le cellule sono legate fra loro mediante una rete a maglie relativamente larghe, costituita da grosse molecole organiche (la cosiddetta matrice extracellulare) e mediante punti di adesione fra le membrane plasmatiche (giunzioni cellulari).
Si classificano tre tipi di giunzioni cellulari: giunzioni ancoranti, giunzioni comunicanti e giunzioni occludenti.
a) Le giunzioni ancoranti (o giunzioni adesive) provvedono a mantenere le cellule in posizioni fisse all'interno dei tessuti tramite connessioni proteiche. Ne esistono di tre tipi: i desmosomi, le giunzioni intermedie e le giunzioni settate. Queste ultime sono tipiche degli invertebrati.
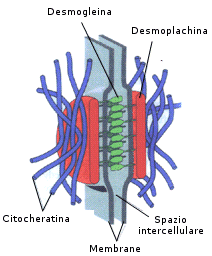
I desmosomi (chiamati anche macula adherens) sono in genere strutture di forma circolare o ellittica. Nella regione contenente un desmosoma è possibile osservare uno spesso strato di materiale denso, la placca, posta immediatamente al di sotto della membrana plasmatica sui due versanti della giunzione. A partire dalla placca, e per una certa distanza in direzione del citoplasma sottostante, si dipartono strutture citoscheletriche ben definite, i filamenti intermedi, detti tonofilamenti, che fungono da ancora citoplasmatica per la giunzione. A seconda del tipo di cellula, essi possono essere costituiti da citocheratina o da vimentina. Le membrane plasmatiche delle cellule che hanno in comune un desmosoma sono tra loro parallele, più o meno allineate e separate da uno spazio di ampiezza costante. Lo spazio extracellulare che le separa è occupato da un fitto intreccio di filamenti sottili e di materiale granulare che va sotto il nome di nucleo del desmosoma. I desmosomi di animali e tessuti diversi contengono proteine rappresentative che sono riscontrabili in modo costante. Tra queste, due delle più grandi, le desmoplachine I e II sono molecole strutturalmente correlate e risultano essere associate alle placche. Un altro gruppo di glicoproteine, le desmogleine, che sono membri della famiglia delle caderine , fungono da ponte citoplasmatico tra le placche ed il materiale extracellulare. Negli animali pluricellulari, i desmosomi congiungono molti tipi diversi di cellule. Queste strutture sono particolarmente abbondanti nei tessuti che sono comunemente sottoposti a stress meccanici in senso laterale o da stiramento, come le cellule dell'epidermide e quelle degli epiteli che rivestono la superficie interna delle cavità corporee.
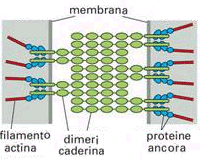
Le giunzione intermedie (o giunzioni aderenti o zonulae adherens), congiungono le cellule di alcuni tessuti animali, come ad esempio quelle del muscolo cardiaco e delle membrane che avvolgono gli organi e che rivestono le cavità corporee. Come nel caso dei desmosomi, anche nella giunzione aderente la regione sottostante la membrana plasmatica è ispessita e dà origine ad una placca. Le placche delle giunzioni aderenti non hanno l'aspetto di uno strato denso e compatto ma piuttosto quello di un reticolo a struttura irregolare. Dalle placche si dipartono in direzione del citoplasma sottostante dei microfilamenti di actina che ancorano la giunzione al citoscheletro. Le placche contengono elevate concentrazioni di vinculina e di altre proteine che normalmente hanno un ruolo di collegamento tra elementi presenti nella membrana plasmatica ed i microfilamenti di actina. Le due placche si saldano nello spazio intercellulare attraverso una maglia proteica di caderina.
Le giunzioni settate (da septum = parete) hanno questo nome in quanto, in sezione trasversale, le membrane che costituiscono le giunzioni risultano essere collegate, nella regione extracellulare, da segmenti trasversali, o setti, a spaziatura regolare. Lo spazio che separa le membrane è caratterizzato da una distanza estremamente regolare, variabile da 15 a 17 nm, dovuta alla presenza di segmenti densi che si alternano a zone chiare, per tutta la lunghezza della giunzione. Tutte le giunzioni settate sono dotate, nella regione extracellulare, di questi segmenti trasversali caratteristici, ma la morfologia dettagliata dei setti, le loro dimensioni e le distanze di spaziatura assumono valori caratteristici in ciascun phylum degli invertebrati.
b) Le giunzioni comunicanti (o giunzioni serrate o gap junction) costituiscono un passaggio aperto attraverso cui ioni e piccole molecole possono direttamente passare da una cellula all'altra. In questo tipo di giunzioni, le membrane plasmatiche delle due cellule adiacenti sono separate da uno spazio molto regolare di 2,7 nm. Il termine usato in inglese, gap, sta a indicare questa separazione sorprendentemente regolare. In queste strutture è stata messa in evidenza la presenza di un grande numero di cilindri cavi disposti con l'asse maggiore perpendicolare alla superficie delle cellule che attraversano lo spazio intercellulare delle giunzioni comunicanti.
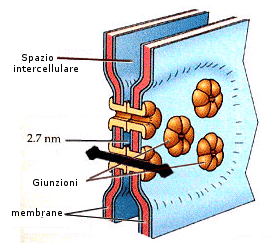
Nei vertebrati, lo stretto canale che si trova al centro del cilindro ha un diametro di circa 1,5 nm nel punto più stretto, dimensione sufficiente a permettere il libero transito di ioni e di piccole molecole. Nelle giunzioni i cilindri appartenenti a due membrane plasmatiche adiacenti si incontrano coda contro coda, e costituiscono un passaggio diretto tra una cellula e l'altra, ma isolano la giunzione rispetto ai liquidi extracellulari. Ogni cilindro della giunzione, definito connessone, è costituito da un numero variabile da 4 a 6 molecole di una proteina detta connessina. A seconda del tipo di cellula dal quale vengono isolate, il peso molecolare delle connessine varia. Le giunzioni comunicanti di un determinato tipo di tessuto possono contenere un unico tipo di connessina o una miscela di due o più tipi diversi. Nonostante le differenze di dimensioni, tutte le connessine hanno una struttura analoga che comprende quattro regioni transmembraniche ad alfa-elica. L'apertura e la chiusura dei canali è regolata dalle cellule. Il sottile strato tra le due cellule impedisce il passaggio di grosse molecole. Sono presenti nel muscolo cardiaco e nei tessuti muscolari lisci.

c) Le giunzioni occludenti (o giunzioni strette o tight junction) hanno il compito di chiudere gli spazi intercellulari alla diffusione, in quanto formano una specie di barriera che impedisce il flusso diretto di molecole e di ioni da una cellula all'altra. Le due membrane cellulari sono strettamente cucite insieme da una proteina di membrana (claudina). Sono le giunzioni più forti perché, oltre a solidità meccanica, impediscono il passaggio di acqua e soluti tra le cellule, formando un sigillo che impedisce il passaggio di sostanze fra il dominio extra-cellulare apicale e il dominio basolaterale. Sono poste ad esempio tra le cellule che rivestono l'apparato digerente, impedendo agli acidi ed agli enzimi di danneggiare i tessuti sottostanti.
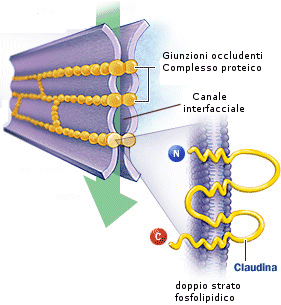
Spesso le giunzioni si trovano coordinate in complessi giunzionali. Un singolo complesso giunzionale è formato da una giunzione occludente, da una giunzione intermedia, da un desmosoma e da una giunzione serrata.
Nelle cellule animali, la membrana plasmatica non presenta generalmente strati esterni di rivestimento. Nei batteri e nei vegetali, invece, all'esterno della membrana si trova una parete rigida, alquanto spessa e robusta, costituita da polisaccaridi complessi (nel caso delle piante superiori, soprattutto da cellulosa). Tale struttura nei batteri ha soprattutto una funzione protettiva; nei vegetali, oltre a questa funzione, la parete svolge un ruolo di sostegno e serve a mantenere la forma tipica della cellula. La parete limita i movimenti della cellula, come pure l'ingresso e la fuoriuscita di materiali.
L'intero volume della cellula, con esclusione del nucleo, è occupato dal citoplasma. Questo comprende una soluzione acquosa concentrata, denominata citosol, nella quale si trovano sospesi gli organuli cellulari.
Il citosol è un gel acquoso, contenente molecole di varie dimensioni. Costituisce il sito di molte funzioni importanti, come la demolizione delle molecole e la sintesi di numerose macromolecole che sono le unità costitutive della cellula. Molte molecole presenti nel citosol sono libere di muoversi per tutte le regioni della cellula; altre hanno una minore libertà di movimento, poiché fanno parte di strutture ordinate, gli organuli, che determinano nella cellula una compartimentazione utile allo svolgimento delle reazioni metaboliche.
Organuli della cellula eucariote
Nel citoplasma troviamo diversi organuli cellulari i quali possono essere raggruppati in sistemi di membrana, mitocondri, ribosomi, sistema citoscheletrico
Sistemi di membrane
Il citosol è invaso da una complessa rete di membrane e vescicole membranose. Costituiscono il sistema interno di membrane (endomembrane) della cellula eucariote: il reticolo endoplasmatico, l'apparato di Golgi, i lisosomi ed i perossisomi
Il reticolo endoplasmatico (RE) è il sistema di membrane più esteso della cellula ed è costituito da una fitta rete di cisterne e tubuli tra loro interconnessi che parte ed è collegato alla membrana nucleare.
Grazie alla presenza sulle sue membrane di proteine enzimatiche il Reticolo è sede di una intensa attività metabolica. La struttura fittamente ripiegata del Reticolo consente alla cellula di avere a disposizione una elevata superficie di lavoro e di tenere separati i diversi processi metabolici. Si distingue un Reticolo endoplasmatico liscio (REL), privo di ribosomi ancorati alle sue membrane ed un Reticolo endoplasmatico rugoso o ruvido (RER) caratterizzato da membrane cosparse di ribosomi sulla faccia citosolica.

il Reticolo endoplasmatico rugoso è costituito da un sistema di sacculi appiattiti impilati, detti cisterne, tra loro connesse. I ribosomi adesi al RER sono responsabili della sintesi delle proteine destinate a far parte delle membrane o ad essere secrete. Inoltre le proteine prodotte vengono ulteriormente rielaborate, con formazione di strutture quaternarie multimeriche o glicosilate (aggiunta di oligosaccaridi) con formazione di glicoproteine. Alla fine di tali processi le molecole prodotte vengono espulse del RER attraverso vescicole raggiungendo i siti di competenza intracellulare (tipicamente l'apparato di Golgi) o extracellulare. Il RER è particolarmente abbondante nelle cellule che presentano attività secretrice (pancreas).
Il Reticolo endoplasmatico liscio si struttura in forma di tubuli e canali ricurvi. ha come compito quello di detossificare sostanze altrimenti dannose per l'organismo, come ad esempio l'etanolo contenuto nelle bevande alcoliche. Per questo motivo è molto diffuso nelle cellule epatiche. Il REL inoltre è sede del metabolismo di fosfolipidi, acidi grassi, steroidi ed è un sito di deposito degli ioni Ca2+. Il REL è infine in grado di degradare gli organuli cellulari per rinnovarli avvolgendoli nelle sue membrane e generando una vescicola di autofagocitosi (o autofagosoma) il cui contenuto verrà successivamente digerito.
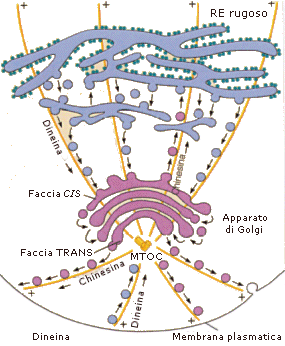
L'Apparato di Golgi (o dittiosoma) si trova tra il reticolo endoplasmatico e la membrana cellulare (da 10 a 20 dittiosomi per cellula). Esso è costituito da una pila di cisterne schiacciate (da 4 a 8 cisterne per dittiosoma), curvate a scodella, con i bordi leggermente rigonfi. Ciascuna cisterna è indipendente dalle altre ed è costituita da una struttura chiusa delimitata da una singola membrana ininterrotta.
L'apparato di Golgi è polarizzato. Il lato rivolto verso il reticolo è detto faccia cis (o CGN = Cis Golgi Network), mentre il lato rivolto verso la membrana è detto faccia trans (o TGN = Trans Golgi Network).
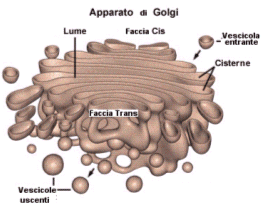
Le proteine che sono state sintetizzate e modificate nel Reticolo endoplasmatico, e da questo rilasciate in vescicole, entrano nel Golgi fondendosi con la faccia Cis. Nel Golgi subiscono ulteriori modifiche mentre sono trasportate verso la faccia trans. Le modalità di migrazione delle sostanze dalla faccia cis verso la faccia trans non sono ancora completamente chiarite.
Secondo l'ipotesi delle "cisterne stazionarie", ogni cisterna possiede il suo corredo enzimatico e dopo aver modificato le sostanze in entrata le cedono, generando vescicole che si fondono con la cisterna successiva.
Secondo l'ipotesi della "maturazione delle cisterne" ogni cisterna modifica gradualmente il suo corredo enzimatico trasformandosi da cisterna cis in cisterna trans ed andando a sostituire la cisterna successiva.
La cisterna trans, completata la trasformazione delle sostanze, le rilascia attraverso vescicole che si formano dalla sua membrana per gemmazione. Le vescicole generate dal Golgi possono essere utilizzate all'interno della cellula, in genere per processi digestivi con formazione di lisosomi o possono essere secrete esternamente (vescicole di esocitosi).
Il lisosoma è una vescicola contenente enzimi idrolitici che si attivano a pH 5. È formato dalla fusione di vescicole idrolasiche del Golgi con vescicole di endocitosi (endosomi). Nelle vescicole idrolasiche del Golgi (o lisosomi primari) sono già presenti gli enzimi litici, ma il pH non è sufficientemente acido perché questi si attivino. Quando si fondono con gli endosomi, i quali sono dotati di pompe protoniche sulla propria membrana in grado di abbassare il pH, gli enzimi vengono attivati e si forma il vero e proprio lisosoma. Esso è ricco di enzimi digestivi in grado di degradare molecole e strutture presenti all'interno della cellula nei loro costituenti elementari per poi venire riutilizzati in altro modo o essere espulsi.
Il lisosoma può degradare macromolecole come proteine, lipidi e carboidrati o anche molecole estranee ingerite dalla cellula. Attraverso questo stesso processo i globuli bianchi sono in grado di digerire i microrganismi patogeni o cellule morte precedentemente fagocitate. La membrana del lisosoma contiene proteine di trasporto per esportare nel citosol i prodotti della digestione e grandi quantità di glucidi legati a lipidi o a proteine della faccia non citosolica come protezione contro i propri enzimi.
I perossisomi sono organuli vescicolari di circa 0,5-1 μm di diametro, delimitati da una singola membrana, presenti in tutte le cellule eucarioti, ma particolarmente abbondanti nelle cellule renali ed epatiche. Svolgono diverse funzioni metaboliche, la più rappresentativa delle quali è senz'altro l'attività ossidativa (respiratoria) a carico di substrati di tipo diverso (soprattutto acidi grassi, ma anche sostanze nocive come l'etanolo con conseguente azione detossificante), che avviene in presenza di una famiglia di enzimi noti come ossidasi. L'attività delle ossidasi avviene con consumo di ossigeno molecolare (O2) e formazione di perossido di idrogeno (o acqua ossigenata H2O2). Il perossido di idrogeno è una molecola altamente reattiva che può facilmente produrre radicali liberi tossici e che i perossisomi rimuovono attivamente usando l'enzima catalasi attraverso la seguente reazione
2H2O2 → O2 + 2H2O
In generale i perossisomi sono in grado di preservare le strutture cellulari dall'azione nociva del perossido di idrogeno (l'acqua ossigenata può essere utilizzata come disinfettante contro i batteri essendo questi privi di perossisomi).
L'appartenenza dei perossisomi al sistema interno di membrane è in realtà ancora oggetto di discussione. I nuovi perossisomi non si formano infatti per scissione dalle membrane del Reticolo o del Golgi, ma si ingrandiscono e si scindono in modo analogo a quanto fanno i mitocondri ed i cloroplasti. Per questo motivo è stata proposta anche per essi (come per i mitocondri ed i cloroplasti) una origine endosimbiontica. Dell'ipotesi endosimbiontica diremo più dettagliatamente quando descriveremo il mitocondrio. Tuttavia, in sintesi, essa ipotizza che alcuni organuli cellulari possano essere antichi microrganismi fagocitati dalla cellula eucariote primordiale in fase di evoluzione che poi stabilì con essi un rapporto di simbiosi. Recentemente è stato tuttavia dimostrato che cellule private dei perossisomi possono generarne di nuovi a partire dal Reticolo. Questa ed altre osservazioni portano oggi a considerare meno probabile l'origine endosimbiontica dei perossisomi a favore di una origine dal sistema interno di membrane.
Il sistema citoscheletrico è costituito dal citoscheletro e da strutture di derivazione citoscheletrica quali ciglia, flagelli, fuso mitotico e dai relativi centri di organizzazione di tali strutture quali centrioli e corpi basali.
Il citoscheletro è un sistema di filamenti proteici presente nel citosol di tutte le cellule animali e vegetali. È costituito da tre tipi principali di filamenti proteici: microtubuli, microfilamenti e filamenti intermedi, connessi sia tra di loro che con altre strutture cellulari grazie a numerose proteine accessorie. Della struttura quaternaria dei microtubuli (tubulina) e dei microfilamenti (actina) abbiamo già detto (vedi capitolo sulle proteine)
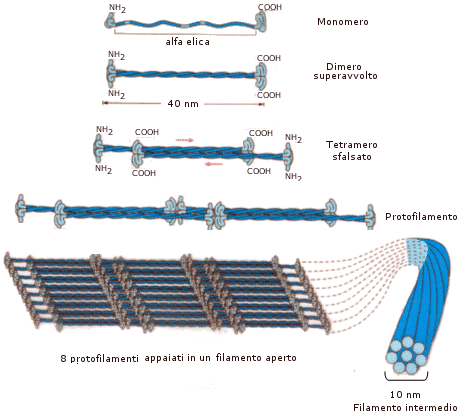
I filamenti intermedi presentano un diametro di circa 10 nm, intermedio appunto tra quello dei microfilamenti (circa 5 nm) e dei microtubuli (circa 25 nm). A seconda del tipo di cellule in cui si trovano possono essere costituiti da proteine di tipo diverso: citocheratina nelle cellule epiteliali, desmina nelle cellule muscolari, neurofilamenti negli assoni delle cellule nervose, vimentina nei fibroblasti del connettivo.
L'elemento di base è un dimero formato da due catene ad alfa elica superavvolte (coiled-coil). I dimeri si appaiano in modo sfalsato ed antiparallelo (estremità ammino-terminale contro estremità carbossi-terminale) a formare tetrametri. I tetrametri si succedono a formare un protofilamento. 2 protofilamenti appaiati formano una protofibrilla. 8 protofilamenti (o 4 protofibrille) si avvolgono a spirale a formare un filamento intermedio
Nelle cellule animali, che mancano di una parete cellulare rigida, il citoscheletro ha un'importanza particolare, in quanto contribuisce a mantenere la struttura e la forma della cellula. Il citoscheletro fornisce infatti un'impalcatura per l'organizzazione interna della cellula e un punto di ancoraggio per organuli ed enzimi.
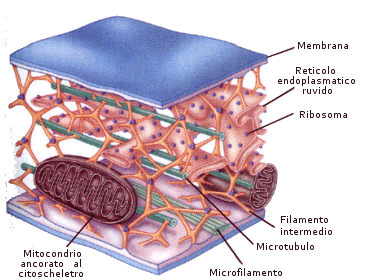
Il citoscheletro è una struttura dinamica, che viene continuamente scomposta e riassemblata. In questo modo è in grado di supportare i movimenti sia dell'intera cellula (pseudopodi, ciglia e flagelli) che degli organuli al suo interno.
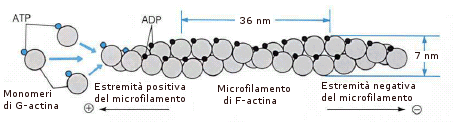
Durante la formazione degli pseudopodi, ad esempio, la cellula costruisce dei microfilamenti nella direzione di avanzamento dell'estroflessione, per dirigerne e sostenerne la crescita. I microfilamenti di actina sono polarizzati (estremità + e -) e si riconoscono non solo da un punto di vista biochimico, ma anche morfologico, perché l'estremità + è appuntita, quella - è sfrangiata. Entrambe le estremità sono siti di crescita con velocità diverse: maggiore nella estremità + e inferiore nella estremità -. La porzione + del filamento di actina rappresenta la zona di attiva crescita e per questo necessita di energia sotto forma di ATP. Se nell'estremità +, i monomeri si legano velocemente, nell'estremità - vengono sottratti monomeri di G-actina che, caricati con ATP si spostano all'estremità positiva per essere assemblati: il risultato è che il filamento, mentre cresce, si sposta. Questo processo viene usato per la formazione di pseudopodi. Quindi, l'avanzamento di un filamento può essere fatto non solo con i monomeri ex-novo, ma anche con unità che derivano dal decremento dell'estremità negativa.
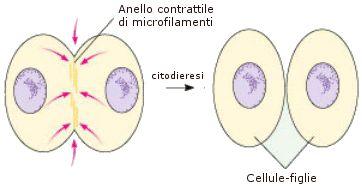
L'actina interviene anche durante la fase finale della divisione cellulare (citodieresi). La cellula eucariote animale si scinde nelle due cellule-figlie grazie ad un anello contrattile di actina (e di miosina, l'actina manifesta proprietà contrattili in presenza di miosina), posto appena sotto la membrana che strozza la cellula-madre nella regione equatoriale.
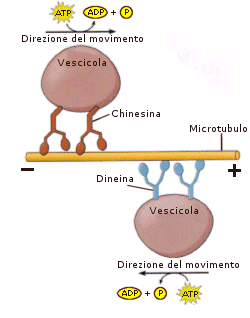
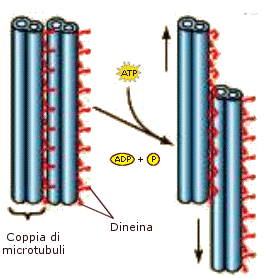
I microtubuli, come i microfilamenti, sono una componente del sistema citoscheletrico fondamentale per i movimenti cellulari. Essi costituiscono infatti una rete interna di binari sui quali possono spostarsi gli organuli e le vescicole cellulari grazie alla presenza di molecole di chinesina e dineina, proteine che fungono da "carrelli trasportatori", trasformando l'energia derivante dall'idrolisi di ATP in energia motrice Inoltre, sempre grazie alla dineina, microtubuli, disposti parallelamente, possono scorrere uno rispetto all'altro consentendo il movimento di ciglia e flagelli. Dineina e chinesina sono una classe di proteine associate ai microtubuli o MAP (Microtubule associated proteins). Dineina e chinesina sono MAP motorie, motori proteici, così chiamate perché usano ATP per guidare il trasporto di vescicole ed organelli o per creare forze di scorrimento tra i microtubuli.
Anche i microtubuli, come i microfilamenti, sono polari , in quanto presentano un'estremità positiva (+), dove la crescita avviene velocemente ed una estremità negativa (-) che influisce poco sull'accrescimento. Le chinesine si muovono verso l'estremità (+) del microtubulo, mentre le dineine verso l'estremità (-).
Nella cellula eucariote animale i microtubuli si formano a partire da un centro organizzatore dei microtubuli o COMT (o MTOC, Microtubule-organizing center) che possiede un punto di innesco per la crescita dei microtubuli costituito da una molecola di tubulina-γ a forma di anello. I microtubuli rivolgono sempre al MTOC la loro estremità (-) e si accrescono allontanandosi da esso con la loro estremità (+). Esistono due MTOC nella cellula animale (la cellula vegetale ne è sprovvista): i centrioli ed il corpo basale.
I centrioli sono due piccoli organuli a cilindretto disposti perpendicolarmente l'uno rispetto all'altro, situati vicino alla membrana nucleare e circondati da un materiale proteico amorfo a formare una struttura denominata centrosoma. I centrioli sono formati da 9 triplette di microtubuli saldate tra loro da filamenti proteici.

Questa struttura è comune anche ai corpi basali. Ogni tripletta di un MTOC presenta la seguente struttura

Quando la cellula non è in fase di divisione cellulare (interfase) la funzione dei centrioli è quella di organizzare la componente microtubulare del citoscheletro. In ogni cellula il movimento delle vescicole non è caotico. Esse si muovono in maniera controllata lungo binari di microtubuli.
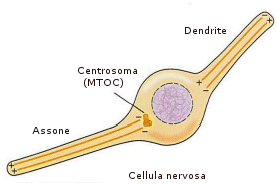
Nel neurone questi binari assumono un significato funzionale cruciale. Le vescicole di neurotrasmettitori costruite nel corpo cellulare vengono trasportate lungo l'assone al bottone terminale (dove trasmettono segnali ad altre cellule) proprio camminando lungo questi microtubuli.
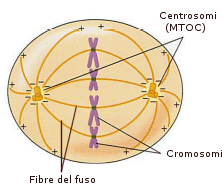 Quando la cellula è in fase di divisione cellulare (mitosi) la funzione
dei centrioli è quella di organizzare il fuso mitotico, un fascio di
microtubuli che fungono da binari per la ordinata e corretta divisione del
materiale genetico (DNA sottoforma di cromosomi) alle due cellule figlie. I due
centrioli si duplicano e migrano ai poli opposti della cellula tendendo fra di
essi un fascio di microtubuli ai quali successivamente aderiranno i cromosomi.
Quando la cellula è in fase di divisione cellulare (mitosi) la funzione
dei centrioli è quella di organizzare il fuso mitotico, un fascio di
microtubuli che fungono da binari per la ordinata e corretta divisione del
materiale genetico (DNA sottoforma di cromosomi) alle due cellule figlie. I due
centrioli si duplicano e migrano ai poli opposti della cellula tendendo fra di
essi un fascio di microtubuli ai quali successivamente aderiranno i cromosomi.
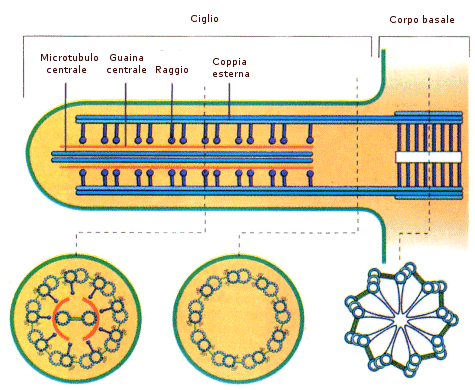
Il corpo basale è un MTOC strutturalmente e funzionalmente simile ad un centriolo. La sua funzione è quella di assemblare e coordinare i microtubuli che formano il citoscheletro di ciglia e flagelli.
Ciglia e flagelli sono estroflessioni filiformi e flessibili della membrana plasmatica, simili a peli, presenti sulla superficie di alcune cellule animali (ciglia e flagelli sono assenti nella cellula eucariote vegetale). Le ciglia sono generalmente corte e numerose sulla superficie cellulare, mentre i flagelli sono più lunghi e presenti nel numero di uno o due per cellula.
La struttura citoscheletrica interna (assonema) di ciglia e flagelli è identica ed è costituita da 9 coppie di microtubuli disposti ad anello attorno a due microtubuli non accoppiati al centro. Questa struttura, detta 9+2, si ritrova in quasi tutte le forme di ciglia e flagelli eucariotici, dai protozoi all'uomo e funziona da motore del movimento. Le coppie esterne di microtubuli sono collegate tra loro da due braccia di dineina e da connessioni proteiche di nexina.
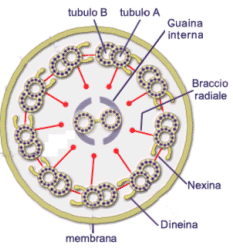
Ogni coppia periferica di microtubuli presenta la seguente struttura

Ciglia e flagelli si flettono dando luogo a un battito regolare come conseguenza dell'energia che si libera quando la dineina idrolizza l'ATP, promuovendo lo scorrimento reciproco dei microtubuli.
L'assonema si connette al corpo basale che penetra nel citosol come una radice e che, come abbiamo visto, presenta la medesima struttura a 9 triplette di microtubuli di un centriolo.
Poiché l'assonema è rigidamente ancorato alla base al corpo basale, lo scorrimento reciproco delle coppie di microtubuli una rispetto all'altra genera la curvatura dell'assonema e quindi il movimento.
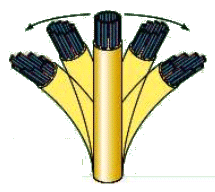
Le ciglia si muovono con un movimento simile a quello delle braccia di un nuotatore

Il flagello si muove con un movimento ondulatorio simile al colpo di una frusta

Negli organismi unicellulari ciglia e flagelli hanno principalmente funzioni di locomozione, permettendo alla cellula di "nuotare" all'interno di un substrato liquido (protozoi ciliati e flagellati). In alcuni casi possono essere utilizzati con funzioni trofiche per creare correnti nel liquido intorno alla cellula in modo da "aspirare" ed indirizzare il cibo verso il luogo in cui verrà digerito, come succede per esempio nelle vorticelle.
Negli organismi pluricellulari sono presenti epiteli ciliati che rivestono organi cavi. Le cilia che rivestono la trachea, ad esempio, hanno una funzione "spazzina". Intercettano le particelle di impurità sospese nell'aria che inspiriamo e, attraverso un ritmico e coordinato movimento verso l'alto (tipo quello della "ola" in uno stadio) le sospingono verso l'imboccatura dell'esofago, dove vengono deglutite.

Nell'orecchio esistono epiteli ciliati con funzione di trasduzione di segnali. Nella coclea, ad esempio, le ciglia che rivestono l'epitelio interno, messe in movimento dalle vibrazioni sonore, stimolano le terminazioni nervose i cui impulsi vengono interpretati dal cervello come suoni.
I ribosomi sono granuli scuri presenti nel citosol oppure sulla membrana nucleare e sul reticolo endoplasmatico rugoso. Sono composti da RNA ribosomiale e materiale proteico e sintetizzano le proteine. La loro disposizione all'interno della cellula è collegata alla funzione di quest'ultima. Se la cellula secerne le proteine prodotte possiede solo ribosomi attaccati al reticolo endoplasmatico (che occupa gran parte del citosol) e alla membrana nucleare. Se la cellula immagazzina le proteine che sintetizza possiede ribosomi liberi. Sono definiti 'liberi' i ribosomi che si trovano nel citoplasma cellulare o che fanno parte dei mitocondri o dei cloroplasti.
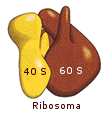
Un ribosoma batterico ha un coefficiente di sedimentazione di 70 S ed è costituito da una subunità grande di 50 S ed una subunità piccola di 30 S.
Lo svedberg (simbolo S) è un'unità di misura del tasso di sedimentazione. Uno svedberg è pari a 10-13 secondi.
L'unità prende il nome dal quello del fisico e chimico svedese Theodor Svedberg, vincitore del Premio Nobel per la chimica nel per il suo lavoro sulla chimica dei colloidi e l'invenzione dell'ultracentrifuga.
Nell'ultracentrifugazione, il tasso di sedimentazione di una particolare macromolecola è calcolato dividendo la velocità di sedimentazione costante (espressa in m/s) per l'accelerazione applicata (espressa in m/s2) e moltiplicando poi per 1013
Il ribosoma della cellula eucariote, invece
(fatta eccezione per quelli contenuti nei mitocondri e nei cloroplasti), è più
grande ed ha un coefficiente di sedimentazione di 80 S. Anch'esso è composto da
due subunità (60 S e 40 S) che nel complesso presentano più di 80 proteine.
Le due subunità del ribosoma sono sintetizzate in una regione interna al nucleo
detta nucleolo. Migrano poi nel citoplasma
attraverso i pori della membrana nucleare e successivamente si uniscono tra
loro ed operano insieme per tradurre un RNA messaggero in una catena
polipeptidica durante la sintesi proteica.
Un mitocondrio è un organulo cellulare di forma allungata, presente in tutti gli Eucarioti (con alcune eccezioni). I mitocondri sono gli organelli addetti alla respirazione cellulare. Nei procarioti sono sostituiti da sacchette contenenti enzimi respiratori. Sono costituiti da due membrane: la membrana interna e la membrane esterna. Lo spazio fra queste due membrane è detto spazio intermembrana. Lo spazio delimitato dalla membrana interna è detto matrice mitocondriale. La membrana interna si estende nella matrice formando delle pieghe dette creste mitocondriali, dove si concentrano gli enzimi respiratori. La membrana mitocondriale esterna è simile a quella del Reticolo Endoplasmatico ed è piuttosto permeabile grazie alla presenza di canali proteici (porine), mentre quella interna è simile alla membrana plasmatica dei batteri, è quasi del tutto priva di colesterolo e presenta trasportatori di membrana altamente selettivi che la rendono assai poco permeabile.
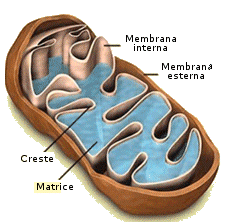
Dopo una breve fase di accrescimento, i mitocondri si dividono per scissione in mitocondri più piccoli. Una volta invecchiati sono eliminati mediante autofagia. Vengono inglobati nella membrana del REL e poi degradati ad opera degli enzimi litici lisosomiali. I mitocondri possiedono un DNA proprio sotto forma di molecole circolari, non legate a proteine, agganciate alla membrana delle creste mitocondriali.
Il mitocondrio è in grado di svolgere molteplici funzioni. La più importante tra esse consiste tuttavia nell'estrarre energia dai substrati organici (respirazione cellulare) per produrre un gradiente ionico che viene sfruttato per sintetizzare ATP. Nel mitocondrio si completano le ultime due fasi della respirazione cellulare.
Con il termine respirazione solitamente si intende il processo fisiologico macroscopico che consiste nella assunzione di O2 e nel rilascio di CO2 da parte di organismi pluricellulari. In biochimica si usa il termine respirazione in senso microscopico per riferirsi ai processi molecolari che implicano consumo di O2 e formazione di CO2 da parte della cellula. Si tratta di una reazione esoergonica di ossidoriduzione in cui un substrato energetico (tipicamente il glucosio C6H12O6) viene ossidato ad anidride carbonica (CO2) cedendo i suoi elettroni all'ossigeno (O2) che si riduce ad acqua (H2O)
C6H12O6 + 6 O2 → 6 CO2 + 6H2O + Energia
La reazione globale è quindi una ossidazione completa del glucosio come in una normale combustione. Tuttavia solo una parte dell'energia liberata viene trasformata in energia chimica sotto forma di ATP. In totale si ottengono 38 ATP per ogni molecola di glucosio. Si ottengono quindi 38 x 7.3 = 277 Kcal/mol. L'efficienza del processo è di circa il 40%. Infatti la reazione di combustione libera 686 Kcal/mol (277/686 ≈ 0.40)
La respirazione cellulare aerobica può essere schematicamente riassunta in tre stadi principali: glicolisi, ciclo di Krebs e fosforilazione ossidativa. Il primo stadio avviene nel citoplasma, gli ultimi due nei mitocondri: Il ciclo di Krebs nella matrice, la fosforilazione ossidativa a livello della membrana interna (creste).
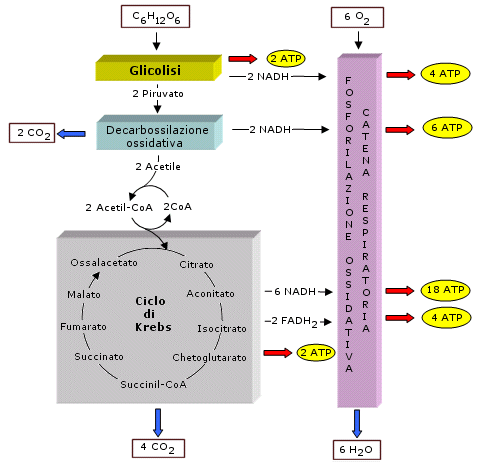
Il temine glicolisi (si pronuncia glicòlisi) deriva da due parole greche che significano "scissione dello zucchero". La glicolisi è una via metabolica pressoché universale dato che è presente non solo negli animali e nelle piante, ma anche nella maggior parte dei microrganismi. La sua universalità e il fatto di essere anaerobica fanno pensare che sia una via metabolica che si è sviluppata agli inizi della vita sulla Terra in organismi procarioti anaerobi che vivevano quando l'atmosfera era ancora povera di O2. La glicolisi è rimasta poi inalterata durante tutta la storia evolutiva grazie alla sua semplicità ed efficienza. La glicolisi è costituita da una sequenza di 10 reazioni che avvengono nel citoplasma e degradano il glucosio in due molecole di acido piruvico (o piruvato) (CH3-CO-COOH) in assenza di ossigeno producendo 2 ATP e 2 NADH. In realtà la glicolisi carica 4 ATP attraverso un processo noto come fosforilazione a livello del substrato, ma la reazione di glicolisi deve essere inizialmente attivata consumando 2 ATP ed il guadagno netto finale è perciò di 2 ATP.
L'acido piruvico viene successivamente ossidato e decarbossilato (decarbossilazione ossidativa = ossidazione ed eliminazione del gruppo carbossile -COOH sottoforma di una molecola di CO2) con formazione di un gruppo acetile (CH3CO-). L'energia e gli elettroni che si liberano da questo processo vanno a caricare 1 NADH (se teniamo conto che il glucosio si è spezzato in 2 piruvati, la decarbossilazione ossidativa carica 2 NADH).
Il gruppo acetile viene agganciato da un coenzima, noto come coenzima A (CoA), con formazione dell'acetil-CoA. Il compito del CoA è di trasportare il gruppo acetile all'interno del ciclo di Krebs affinché venga completato il processo ossidativo. Si tenga presente che l'acetil-CoA può essere generato anche a partire da altri substrati energetici (tipicamente lipidi).
Il ciclo di Krebs, o ciclo degli acidi tricarbossilici (TCA cycle) o ciclo dell'acido citrico, consiste in una serie di reazioni che avvengono all'interno dei mitocondri, nello spazio della matrice. Queste reazioni sono realizzate attraverso otto tappe enzimatiche e hanno lo scopo di ossidare completamente i due atomi di carbonio del gruppo acetile formando due molecole di CO2. Durante questo processo si libera energia ed elettroni (e ioni H+) che vanno a caricare 3 NADH, 1 FADH2 e 1 ATP. Anche in questo caso l'ATP si forma come conseguenza di un processo di fosforilazione a livello del substrato. Tenendo conto che una molecola di glucosio fornisce al ciclo di Krebs 2 gruppi acetile, il guadagno ottenuto dal ciclo di Krebs per molecola di glucosio sarà di 6 NADH, 2 FADH2 e 2 ATP. Se sommiamo a questo le molecole che si sono caricate in precedenza (2 ATP e 2 NADH per la glicolisi, 2 NADH per la decarbossilazione ossidativa) si ottengono:
10 NADH 2 FADH2 4 ATP
Le ossidoreduttasi (NADH, FADH2) che si sono finora caricate rappresentano degli accettori temporanei di elettroni. Esse devono scaricarsi e cedere elettroni ed energia ad un accettore finale e permanente. Come tutti gli enzimi ed i coenzimi, anche le ossidoreduttasi sono presenti in piccolissime quantità. Se rimanessero bloccate nella loro forma ridotta (alcuni veleni producono questo effetto) le prime due fasi della respirazione cellulare si bloccherebbero completamente, causando la morte dell'organismo.
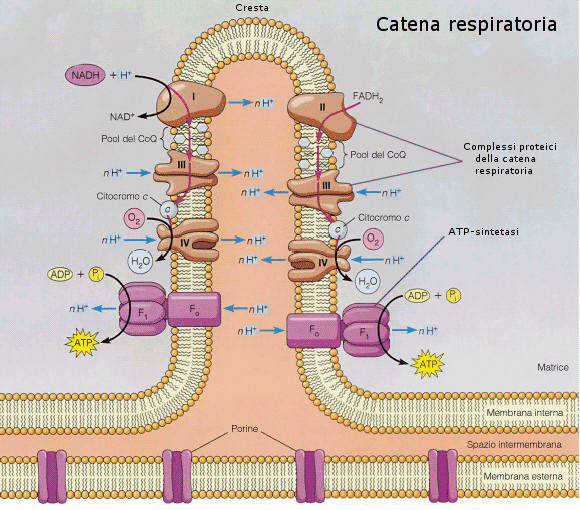
Nel terzo stadio NADH e FADH2 si scaricano, cedendo elettroni (e ioni H+) all'ossigeno che, riducendosi ad acqua, costituisce l'accettore finale e permanente di elettroni. Il trasferimento di elettroni non avviene direttamente, ma attraverso tutta una serie di molecole intermedie (citocromi), attraverso una sorta di catena di trasporto degli elettroni, nota come catena respiratoria. In questo modo l'energia contenuta nelle ossidoreduttasi viene liberata in piccole quantità ad ogni passaggio e può essere utilizzata per caricare ATP.
Ogni NADH che si scarica nella catena respiratoria è in grado di caricare 3 ATP
Ogni FADH2 che si scarica nella catena respiratoria è in grado di caricare 2 ATP
I 10 NADH caricheranno quindi 30 ATP, i 2 FADH2 caricheranno altri 4 ATP, per un totale di 34 ATP che, sommati ai 4 ATP già ottenuti portano il guadagno totale della respirazione aerobica a 38 ATP. In alcune cellule il trasporto all'interno dei mitocondri dei 2 NADH che si erano formati durante la glicolisi richiede un consumo di 2 ATP. In questo caso la resa finale sarà di 36 ATP.
La catena respiratoria è costituita da quattro complessi proteici che contengono gruppi prostetici redox saldamente legati. Gli elettroni vengono trasferiti da un gruppo redox al successivo. Durante il passaggio di elettroni i complessi 1, 3 e 4 sono in grado di generare un flusso di ioni H+ dalla matrice verso lo spazio intermembrana e per questo sono chiamati pompe protoniche.
In questo modo si genera ai due lati della membrana delle creste una differenza di concentrazione in ioni H+ (gradiente elettrochimico). Sotto la spinta di questo gradiente gli ioni H+ rientrano nella matrice utilizzando l'ATP-sintetasi, un enzima transmembranico in grado di convertire il flusso di ioni H+ in energia chimica per la sintesi di ATP (teoria chemiosmotica).
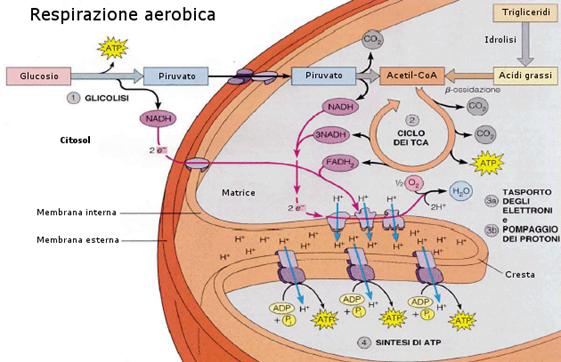
L'intero processo può essere così schematizzato:
Respirazione aerobica - schema
L'evoluzione delle cellule eucarioti ha comportato un notevole salto evolutivo, con il passaggio da cellule procarioti semplici a cellule ben suddivise in parti funzionali, il tutto senza passare per alcuna forma intermedia.
Il rompicapo di questa mancanza di forme è stato spiegato con la teoria dell'endosimbiosi. Sulla base delle forti somiglianze tra i batteri da una parte e i mitocondri e i cloroplasti delle cellule eucarioti dall'altra, sembra probabile che questi organelli si siano originati da procarioti liberi, riparatisi all'interno di cellule eterotrofe più grandi.
I mitocondri si possono ricondurre ad organismi aerobi analoghi ai batteri, mentre i plastidi ad organismi autotrofi simili alle alghe azzurre. Entrambi sarebbero stati assimilati, nel corso dell'evoluzione, in qualità di simbionti da procarioti più grandi. Tali simbionti, perduta la loro autonomia, sono poi divenuti elementi stabili e necessari della cellula eucariote.
Tra le prove a favore di questa teoria ci sono, tra le altre:
il fatto che i mitocondri ed i plastidi si formino solamente per divisione autonoma da organuli uguali.
Il fatto che, come i procarioti, contengano DNA anulare.
Il fatto che la loro membrana esterna sia di tipo eucariotico (membrana di fagocitosi), mentre quella interna di tipo procariotico (membrana originale dell'endosimbionte fagocitato).
Sempre per la stessa teoria, si fanno derivare da batteri analoghi alle spirochete anche le ciglia e i flagelli degli eucarioti con struttura 9+2 (la tipica struttura con 2 microtubuli centrali (singoletto) circondati radialmente da 9 coppie (doppietti) di microtubuli formanti un anello periferico)
Tutti gli eucarioti autotrofi contengono cloroplasti, e tutti gli eucarioti contengono mitocondri: entrambe gli organelli sembra siano stati acquisiti attraverso eventi simbiotici differenti (come si può vedere dall'immagine).
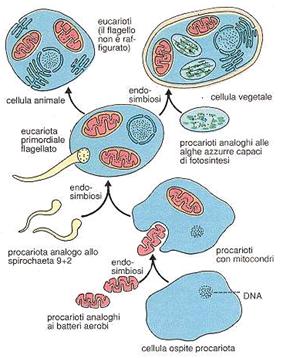
Nella respirazione cellulare aerobica l'accettore finale di elettroni è l'ossigeno. Si definiscono anaerobiche le vie metaboliche di demolizione dei substrati energetici in cui l'accettore finale di elettroni non è l'ossigeno. Si tratta di processi di gran lunga meno efficienti, dal punto di vista energetico della respirazione aerobica, ma sufficienti a garantire la sopravvivenza di molti microrganismi. Ci occuperemo di due esempi importanti di respirazione anaerobica: la fermentazione lattica e la fermentazione alcolica.
Si tratta di due vie metaboliche che utilizzano la glicolisi come tappa iniziale.
Nella fermentazione lattica i due NADH che si formano dal processo glicolitico si scaricano sul piruvato, riducendolo ad acido lattico (CH3-CHOH-COOH). Alla fine del processo il guadagno energetico si riduce quindi ai 2 ATP che si erano formati durante la glicolisi. La fermentazione lattica è alla base della trasformazione del latte in yogurt grazie all'attività di particolari batteri.
La fermentazione lattica può avvenire anche nei nostri muscoli, quando il lavoro muscolare è particolarmente intenso e l'apporto di ossigeno non è sufficiente a soddisfare le richieste energetiche. In queste condizioni la glicolisi può essere accelerata (non richiede ossigeno), ma l'eccesso di piruvato che si forma e che non può essere assorbito dal ciclo di Krebs, viene convertito in acido lattico. La diminuzione di pH (lattacidosi) che si genera nelle cellule muscolari è responsabile dell'indolenzimento muscolare e può provocare crampi.
Anche nella fermentazione alcolica i due NADH che si formano dal processo glicolitico si scaricano sul piruvato. In questo caso il piruvato viene però anche decarbossilato, con liberazione di una molecola di anidride carbonica (CO2) e formazione di etanolo o alcol etilico (CH3-CH2OH). Alla fine del processo il guadagno energetico è sempre di 2 ATP come nella fermentazione lattica. La fermentazione alcolica è alla base della produzione di bevande alcoliche fermentate come il vino, la birra, il sidro il sakè etc ad opera di microrganismi unicellulari fungini, noti come lieviti. La liberazione di CO2 durante questo processo permette di ottenere fermentati gasati (birra, spumante). La fermentazione alcolica viene utilizzata anche nei processi di lievitazione. Durante la lievitazione del pane, ad esempio, i carboidrati contenuti nella farina vengono trasformati in etanolo e anidride carbonica. La CO2 che si libera nell'impasto lo gonfia rendendolo spugnoso, La successiva cottura fa evaporare sia l'etanolo che il gas.
La struttura di maggiori dimensioni all'interno della maggior parte delle cellule vegetali e animali è il nucleo. Il nucleo cellulare contiene il DNA ed ha quindi una funzione essenziale nella trasmissione dei caratteri ereditari (duplicazione DNA) e nel controllo del metabolismo cellulare (trascrizione dei geni e controllo della sintesi proteica). I geni contenuti nel DNA vengono, infatti, copiati all'interno del nucleo in una molecola di acido ribonucleico (trascrizione), chiamata RNA messaggero (mRNA), che passa nel citoplasma e, interagendo con i ribosomi, dirige la formazione di molecole proteiche a partire da singoli amminoacidi veicolati da molecole di RNA di trasporto.
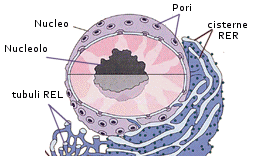 La
forma del nucleo cambia notevolmente, generalmente seguendo la geometria della
cellula, dunque cellule cilindriche avranno nuclei oblunghi, mentre cellule
cubiche avranno nuclei sferici. L'involucro nucleare è composto da due membrane
concentriche, ciascuna di spessore di 8 nm circa, che delimitano il lume della cisterna
perinucleare di 15-40 nm.
La
forma del nucleo cambia notevolmente, generalmente seguendo la geometria della
cellula, dunque cellule cilindriche avranno nuclei oblunghi, mentre cellule
cubiche avranno nuclei sferici. L'involucro nucleare è composto da due membrane
concentriche, ciascuna di spessore di 8 nm circa, che delimitano il lume della cisterna
perinucleare di 15-40 nm.
La membrana nucleare esterna è in continuità con le membrane del reticolo endoplasmatico ruvido e, analogamente, la sua superficie esterna è cosparsa di ribosomi. Lo spazio perinucleare è in continuità con il lume del reticolo endoplasmatico. Le membrane nucleari esterna e interna sono interrotte a livello dei pori dove le due membrane si fondono.
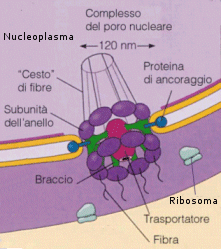
I pori nucleari sono circondati da una struttura ad anello, detta complesso del poro nucleare, formata da otto granuli di origine proteica. Attraverso i pori nucleari avviene il trasporto selettivo di grosse molecole o particelle. Per esempio, tutte le molecole di RNA e i ribosomi della cellula sono sintetizzati nel nucleo e quindi trasferiti nel citosol, mentre tutte le proteine che svolgono le loro funzioni all'interno del nucleo sono sintetizzate nel citosol (unica sede della sintesi proteica) e devono venire introdotte nel nucleo.
All'interno del nucleo si trova il carioplasma (o nucleoplasma), in cui si distingue una parte fluida, detta cariolinfa (o succo nucleare) ed un materiale basofilo in essa disperso che, proprio per la forte affinità con i coloranti (basici) è stato chiamato cromatina. Nel carioplasma sono inoltre presenti corpi sferoidali fortemente basofili (solitamente più scuri del resto del nucleo) a causa dell'abbondanza di RNA, ma affini anche a molti coloranti acidi (al contrario del resto del nucleo) a causa della presenza di proteine basiche, detti nucleoli. La grandezza di un nucleolo va da 1 a 3 μm. Il numero di nucleoli osservabili in un nucleo è compreso tra uno e sei.
Il nucleolo è una regione deputata alla trascrizione dell'RNA ribosomiale e al successivo assemblaggio delle subunità ribosomiali. Queste, una volta sintetizzate, migrano nel citoplasma e costituiscono la sede della sintesi proteica
La cromatina è costituita da DNA e proteine nucleari classificate come istoniche (istoni) e non-istoniche (non-istoni).
La cromatina può manifestarsi in una forma più o meno condensata, come conseguenza del diverso grado di compattazione del DNA attorno agli istoni.
Gli istoni sono proteine relativamente piccole, a forte carica positiva (per la presenza di lisina e arginina). La funzione delle proteine istoniche è quella di organizzare il DNA compattandolo in modo tale da consentire alle cellule di conservarlo in un volume ristretto come quello del nucleo. La famiglia degli istoni comprende le proteine H1, H2A, H2B, H3 e H4.
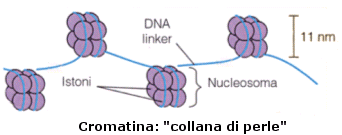
Nel primo livello di compattazione il DNA si avvolge per un giro e tre quarti attorno ad un 'rocchetto' proteico, costituito da 8 istoni (ottamero) a due a due uguali (H2A, H2B, H3 e H4). Questa struttura è nota come nucleosoma. I nucleosomi hanno un diametro di circa 11 nm ed il DNA vi si avvolge intorno per un totale di 146 coppie di nucleotidi. Tra un nucleosoma e l'altro, è presente un filamento di DNA detto linker, che può variare in lunghezza tra poche basi fino a circa 150.
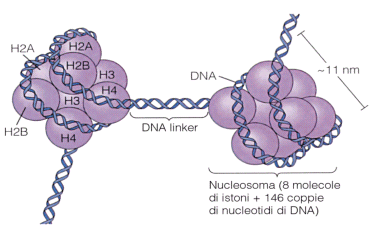
Questo dà alla cromatina l'aspetto di una "collana di perle", osservabile al microscopio elettronico
 In
questo stato la cromatina viene detta decondensata
o despiralizzata ed è nota con il
nome di eucromatina. L'eucromatina è
trascrizionalmente attiva. In altre parole il DNA in questo stato è accessibile
agli enzimi che trascrivono l'informazione genetica e sintetizzano RNA.
In
questo stato la cromatina viene detta decondensata
o despiralizzata ed è nota con il
nome di eucromatina. L'eucromatina è
trascrizionalmente attiva. In altre parole il DNA in questo stato è accessibile
agli enzimi che trascrivono l'informazione genetica e sintetizzano RNA.
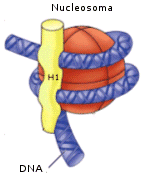
Ogni nucleosoma è associato all'istone H1, il quale sembra abbia la funzione di compattare ulteriormente i nucleosomi.
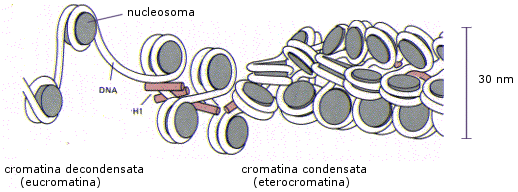
Nel secondo livello di condensazione i nucleosomi, compattati dall'istone H1, formano una fibra da 30 nm di diametro. I livelli di condensazione superiori all'eucromatina sono invece trascrizionalmente inattivi (il DNA non può essere trascritto) e sono noti con il termine eterocromatina.
Le fibre da 30 nm possono ripiegarsi su se stesse in anse, le quali possono infine raggiungere il grado finale di compattazione condensandosi in cromosomi (corpuscoli di cromatina). Le anse sono stabilizzate da una impalcatura (scaffold) di proteine non-istoniche
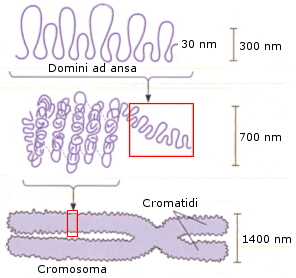
I cromosomi compaiono nel nucleo cellulare solo al momento della divisione cellulare (mitosi e meiosi), quando la cellula-madre si divide in due cellule-figlie. Durante la divisione cellulare infatti la cellula-madre deve fornire alle cellule-figlie una copia del suo DNA e la completa condensazione della cromatina in cromosomi rende ovviamente più agevole l'ordinata ripartizione del materiale genetico.
Il cromosoma presenta una forma bastoncellare costituita da due filamenti identici di DNA, detti cromatidi fratelli, uniti insieme a livello di una strozzatura detta centromero che dà loro una forma ad X. A ciascun centromero è associata una placca proteica, detta cinetocore, che serve come punto di attacco per le fibre del fuso mitotico.
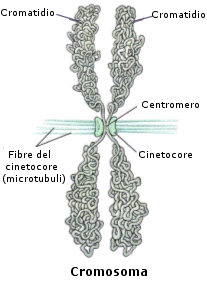
Le estremità di un cromosoma sono dette telomeri.
Il telomero è una struttura "specializzata" presente alle due estremità di ogni cromosoma. Le funzioni fondamentali del telomero sono quelle di mantenere l'integrità delle estremità cromosomiche e di opporsi ad una eccessiva proliferazione cellulare.
Il telomero impedisce infatti la deleteria fusione tra cromosomi, che avviene ad esempio facilmente per unione di due estremità cromosomiche spezzate.
Inoltre il telomero svolge, in condizioni
normali, l'importante funzione di marcatempo, cioè conteggia ( e impone) il
numero di volte in cui una cellula può riprodursi. Da studi su colture di
cellule umane è stato possibile stabilire che le replicazioni cessano dopo
50-70 divisioni cellulari.
Il telomero situato, come già detto, alle due estremità del cromosoma,
strutturalmente è un insieme di brevi sequenze identiche (sequenze ripetitive) di
basi azotate ripetute migliaia di volte (DNA in tandem o DNA satellite).
Queste sequenze ripetute (dette anche terminali per la loro posizione nel
cromosoma) sono prive di geni e non hanno pertanto proprietà codificanti (non
producono proteine).
In tutti i mammiferi, compresa la specie umana, la "firma" molecolare del
telomero, più volte reiterata in successione, consiste di sei lettere riferite
alla sequenza delle basi azotate costituenti il modulo fondamentale: TTAGGG.
La sequenza ripetuta in tandem può essere espressa come: (TTAGGG)n, dove n è il numero delle ripetizioni (il valore medio nella specie umana è circa n = 2000).
Il telomero subisce un accorciamento ogni volta che i cromosomi vengono duplicati nella fase S (S: sintesi) della divisione cellulare (mitosi). L'enzima telomerasi, qualora presente, provvede ad integrare la porzione delle sequenze ripetute che sono state consumate nell'evento replicativo, ripristinando le dimensioni originarie del telomero.
Tuttavia, in condizioni normali la telomerasi
è presente e attiva soltanto nelle cellule germinali (spermatozoi, cellule uovo
e loro cellule progenitrici) e nelle cellule staminali.
Nel corso della vita la telomerasi scompare in quasi tutte le cellule somatiche
(rimane nelle staminali) e pertanto viene a determinarsi un accorciamento
progressivo dei telomeri in rapporto al numero delle replicazioni cellulari nel
frattempo intervenute (orologio mitotico). La regressione telomerica va a
coincidere nell'ultima fase con il fenomeno dell'invecchiamento cellulare
(senescenza replicativa). Pertanto, dopo un numero calcolabile di duplicazioni
(50-70 nelle cellule umane), al di sotto di una certa soglia critica di
riduzione della lunghezza telomerica, la crescita cellulare si blocca, per
passare da una fase di senescenza ad una fase di crisi, nella quale la letalità
cellulare prevale a seguito della fusione e della frammentazione dei cromosomi
(caos genetico) e la cellula entra in apoptosi
(morte cellulare programmata).
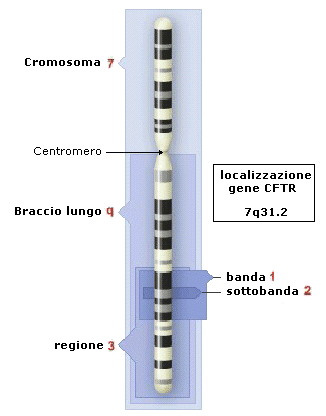 Dal punto di vista genetico, un cromosoma può
essere definito come una sequenza lineare di geni. Al microscopio ottico, i
cromosomi sono distinguibili tra loro per le dimensioni, per la 'forma'
e per la posizione del centromero. Ulteriori distinzioni si possono effettuare
con opportuni trattamenti chimici, che evidenziano un bandeggio (alternanza di
bande trasversali con diversa pigmentazione) che
identificano precise regioni. I cromosomi sono numerati progressivamente in
base alle loro dimensioni: il cromosoma 1 è il più grande di tutti mentre il
cromosoma 22 è il più piccolo. In ogni cromosoma si distinguono un braccio
corto, indicato come "p", un braccio lungo, indicato come "q". Ogni braccio è
suddiviso in regioni e bande, classificate con numeri progressivi a partire dal
centromero
verso l'estremità del braccio corto e del braccio lungo. Ogni
regione è suddivisa in bande ed in sottobande, classificate anch'esse con
numerazione standardizzata a partire dal centromero. Ad esempio il gene della
fibrosi cistica (CFTR - cystic fibrosis transmembrane regulator) è localizzato
nella regione 3 (banda 1, sottobanda 2) del braccio lungo (q) del cromosoma 7
ed è quindi individuato come 7q31.2
Dal punto di vista genetico, un cromosoma può
essere definito come una sequenza lineare di geni. Al microscopio ottico, i
cromosomi sono distinguibili tra loro per le dimensioni, per la 'forma'
e per la posizione del centromero. Ulteriori distinzioni si possono effettuare
con opportuni trattamenti chimici, che evidenziano un bandeggio (alternanza di
bande trasversali con diversa pigmentazione) che
identificano precise regioni. I cromosomi sono numerati progressivamente in
base alle loro dimensioni: il cromosoma 1 è il più grande di tutti mentre il
cromosoma 22 è il più piccolo. In ogni cromosoma si distinguono un braccio
corto, indicato come "p", un braccio lungo, indicato come "q". Ogni braccio è
suddiviso in regioni e bande, classificate con numeri progressivi a partire dal
centromero
verso l'estremità del braccio corto e del braccio lungo. Ogni
regione è suddivisa in bande ed in sottobande, classificate anch'esse con
numerazione standardizzata a partire dal centromero. Ad esempio il gene della
fibrosi cistica (CFTR - cystic fibrosis transmembrane regulator) è localizzato
nella regione 3 (banda 1, sottobanda 2) del braccio lungo (q) del cromosoma 7
ed è quindi individuato come 7q31.2
Le dimensioni dei cromosomi (e del DNA in generale) possono essere espresse in unità 'bp' Una unità bp (base pair) si riferisce ad una singola coppia di basi complementari (A-T o C-G) che si succedono nel filamento di DNA. Si usano ovviamente i multipli di tale unità come:
1 kbp = chilobase = mille basi
1 Mbp = megabase = un milione di basi
1 Gbp = gigabase = un miliardo di basi
Le dimensioni dei cromosomi sono comprese in un intervallo di 50-280 Mbp. Poiché il genoma umano contiene circa 3200 Mpb (3,2 Gbp), la dimensione media dei 23 cromosomi è di circa 140 Mbp (3200/23).

Il numero di geni contenuto nel DNA umano è ancora oggetto di discussione. Attualmente si ritiene che esso sia intorno a 20.000-30.000 geni. Un gene ha una dimensione media di 27 kbp (27.000 basi), di cui solo 1-1,5 kbp (1000 - 1500 basi) sono codificanti per la proteina (esoni), mentre la frazione residua (introni) non viene tradotta ed ha probabilmente funzioni di regolazione dell'espressione genica.
Questo significa che solo il 25% del genoma è costituito da DNA genico, per circa 800 Mbp (30.000 geni x 27 kbp/gene) e solo l'1,5% del genoma va a formare il DNA codificante per circa 45 Mbp (30.000 geni x 1,5 kbp/gene). Il rimanente 75% del genoma costituisce il DNA extragenico (o intergenico)
Dunque circa il 98.5% del genoma umano è composto di sequenze non codificanti. Si definisce DNA non codificante ogni sequenza di DNA in un genoma non soggetta a trascrizione in RNA o rimossa dall'RNA messaggero prima della traduzione (introni) e che viene perciò considerata, allo stato attuale delle conoscenze apparentemente priva di funzione, nonostante diverse ipotesi in merito siano state formulate. Tali regioni di DNA erano indicate con i termini di junk DNA o DNA spazzatura.
Un tipo particolare di DNA non codificante è costituito dalle cosiddette sequenze ripetitive che costituiscono una frazione elevata del genoma (35-50%). Il DNA ripetitivo è costituito da sequenze di DNA che si ripetono uguali molte volte nel genoma. Il DNA ripetitivo viene classificato in DNA in tandem e DNA intersperso,
Il DNA in tandem è noto anche come DNA satellite, poiché, in seguito a centrifugazione del DNA, si concentra in sottili bande satellite distinte dalla banda principale del DNA. Il DNA in tandem costituisce circa il 5% del genoma ed è formato da sequenze molto brevi di basi (10-100 bp) i cui compaiono soprattutto Adenina e Timina, che si ripetono uguali una dopo l'altra (in tandem) per tratti lunghi fino a qualche centinaio di kbp. Si trova localizzato nelle regioni centromeriche e telomeriche del cromosoma
Il DNA intersperso è costituito da sequenze che si ripetono in punti diversi del DNA, disperse in tutto il genoma. Il DNA intersperso è frutto dell'attività dei trasposoni. I trasposoni sono elementi mobili che si trovano nel genoma di tutti gli organismi. Sono in grado, con meccanismi diversi, di saltare da un punto all'altro del genoma. Esistono due tipi di trasposoni: - i trasposoni a DNA che, tramite l'enzima trasposasi, si staccano dal filamento principale sotto forma di un breve filamento anulare di DNA e si inseriscono in un altro punto del filamento principale (trasposizione conservativa)
- i retrotrasposoni che utilizzano un processo di trascrizione inversa (retrotrascrizione), tramite il quale un filamento di RNA sintetizza numerose segmenti identici di DNA che si inseriscono in punti diversi del filamento principale di DNA (trasposizione replicativa). Sono retrotrasposoni le sequenze ripetitive intersperse note come SINE (Short INterspersed Elements = elementi interspersi corti), lunghe 100-300 bp e LINE (Long INterspersed Elements = elementi interspersi lunghi), lunghe 6-8 Kbp. Nell'uomo predominano tra le SINE, la famiglia Alu, e tra le LINE, la famiglia L1.
Spesso le unità di misura delle dimensioni del DNA (kbp, Mbp, Gbp) vengono abbreviate in kb, Mb e Gb, generando non poca confusione con le corrispondenti unità di misura dell'informazione.
Kb è infatti il chilobyte, Mb il megabyte e Gb il gigabyte.
La confusione nasce dal fatto che le sequenze di DNA contengono dell'informazione (genetica) che può dunque essere misurata in bit, byte e suoi multipli (kb, Mb, Gb). Vediamo come.
Il bit (contrazione di binary digit = cifra binaria) è l'unità di misura dell'informazione. Il bit è uno dei due simboli del sistema numerico binario, classicamente chiamati zero (0) ed uno (1) che utilizziamo per individuare univocamente due alternative.
Per individuare in modo univoco le 4 basi (Adenina, Timina, Guanina e Citosina) sono necessari 2 bit (due cifre binarie).
Ad esempio:
Adenina = 00 Timina = 01 Guanina = 10 Citosina = 11
Così se volessimo registrare l'intera informazione contenuta nel genoma umano (3,2 Gbp) sarebbero necessari 6,4 miliardi di bit (6,4 Gbit = 6.400 Mbit).
Tenendo poi presente che, per definizione, 1 byte = 8 bit, il genoma umano contiene informazioni per 6.400 Mbit/8 = 800 megabyte (Mb).
In pratica, per trasformare le dimensioni del DNA, espresse in bp, in quantità di informazione in esso contenuta, espresse in byte, è sufficiente dividere per 4 (si moltiplica per 2 per ottenere i bit e si divide per 8 per ottenere i byte).
Il risultato è certamente sorprendente se si pensa che attualmente (settembre 2006) Windows XP richiede circa 1500 Mb di spazio su disco rigido per gestire un computer, quando con 'soli' 800 Mb il DNA gestisce l'intero nostro organismo.
Numero e struttura dei cromosomi costituiscono il cariotipo, ben evidenziabile (e fotografabile) durante la divisione cellulare.
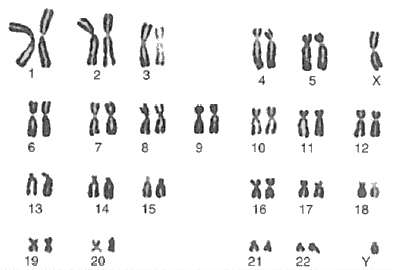
In molte cellule i cromosomi non sono tuttavia uno diverso dall'altro, ma si presentano in coppie morfologicamente simili. I cromosomi di una medesima coppia si dicono omologhi.
Le cellule che hanno coppie di cromosomi omologhi sono dette diploidi (2n), mentre sono definite aploidi (n) le cellule che presentano un'unica serie di cromosomi uno diverso dall'altro.
n è il numero di cromosomi di una singola serie.
Le cellule diploidi sono tipiche degli organismi che si riproducono sessualmente e le due serie di omologhi sono una di provenienza materna e l'altra di provenienza paterna.
Ogni specie presenta un corredo cromosomico caratteristico per forma e numero. Le cellule diploidi umane presentano 23 coppie di omologhi (46 cromosomi) e quindi n = 23.
Nella specie umana la 23ma è la coppia dei cromosomi sessuali, che definisce il sesso genetico dell'individuo. Nella femmina è costituita da due cromosomi omologhi, detti X. Nel maschio invece è costituita da un cromosoma X (come quelli presenti nella femmina e da un cromosoma più piccolo detto Y.
Le proteine nucleari (istoni e non-istoni) non hanno solo funzioni strutturali (compattazione del DNA), ma svolgono un ruolo importante nella regolazione dell'espressione genica, stabilendo quali regioni del DNA debbano essere accessibili all'apparato trascrizionale (il DNA in forma condensata non può infatti essere ne' trascritto ne' duplicato). Nel nucleo avvengono in definitiva vari fenomeni di regolazione e attivazione dei geni, mediati da proteine, in modo che una proteina, codificata da un tratto del DNA (gene), venga prodotta esclusivamente nei tempi e nelle quantità richiesti dalla fisiologia della cellula.
Le proteine non istoniche sono proteine a carica negativa e sono classificate in:
proteine che regolano la trascrizione genica;
enzimi attivi nella trascrizione, replicazione e riparazione del DNA;
proteine che partecipano al mantenimento della struttura della cromatina dallo stato decondensato a quello compatto.
Estratto da ''
Tutte le cellule di un organismo pluricellulare possiedono lo stesso genoma. Tuttavia ogni cellula si specializza nell'eseguire determinate funzioni ed utilizza solo una parte delle informazioni contenute nel suo DNA (esprime solo certe proteine). Per ogni tipo cellulare inoltre, il tipo e la quantità di proteine prodotte dipendono anche dalle necessità che la cellula presenta in un determinato istante. Le cellule devono dunque possedere dei meccanismi che permettano loro di esprimere certi geni e silenziarne altri.
Si distinguono geni costitutivi che sono costantemente attivi (es. geni che codificano per gli enzimi della glicolisi) e geni regolati la cui espressione è regolata in relazione al fabbisogno cellulare (es. sintesi adattativa di enzimi).
I meccanismi di regolazione genica più semplici e meglio compresi sono quelli utilizzati dai procarioti (batteri). I batteri utilizzano sistemi di regolazione genica noti come "operoni".
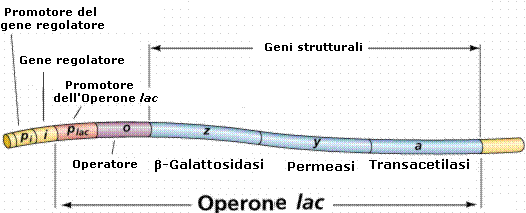
L'operone è una unità funzionale del DNA costituita da un gruppo di geni contigui strettamente correlati, responsabili della sintesi coordinata di una proteina e della regolazione della sintesi stessa.
Un operone contiene:
uno o più geni strutturali destinati ad essere trascritti (sintesi mRNA) e ad esprimere la proteina
un promotore che precede i geni strutturali, costituito da una particolare sequenza di DNA che indica dove debba iniziare la trascrizione. Un promotore occupa una posizione adiacente al gene o al gruppo di geni di cui controlla la trascrizione e la sua funzione è quella di rappresentare una zona di riconoscimento e di attacco per una molecola di RNA polimerasi , l'enzima deputato alla sintesi di RNA messaggero (mRNA) a partire da uno stampo di DNA
un gene operatore che controlla l'espressione genica dei geni strutturali.
L'azione dell'operatore è a sua volta sotto controllo di un gene regolatore che non fa parte dell'operone. Il gene regolatore è in gradi di bloccare o sbloccare il gene operatore in funzione della presenza o meno nella cellula di un particolare metabolita.
L'operone lattosio o operone lac in Escherichia coli è stato il primo operone studiato.
L'operone lattosio produce gli enzimi necessari per l'utilizzazione del lattosio da parte del batterio. Gli enzimi sono codificati da tre geni strutturali adiacenti Z, Y ed A. Questi geni vengono trascritti in un solo mRNA che viene poi tradotto nelle tre proteine. In particolare il gene lacZ codifica per la β-galattosidasi, enzima in grado di idrolizzare il lattosio (un disaccaride) in glucosio e galattosio
|
DNA |
Funzione |
|
Promotore del regolatore (PI) |
Controllo della trascrizione del Regolatore |
|
Regolatore (I) |
Codifica il repressore |
|
Promotore (Plac) |
Controllo della trascrizione dei geni a valle |
|
Operatore (O) |
Bersaglio del repressore |
|
lacZ |
Codifica beta-galattosidasi |
|
lacY |
Codifica lattosio-permeasi |
|
lacA |
Codifica transacetilasi |

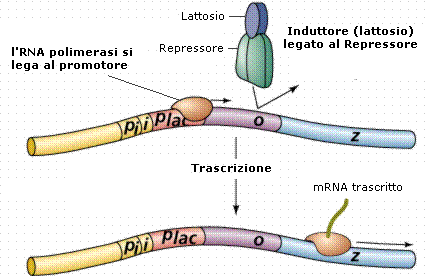
L'operone lac è un esempio di operone inducibile, tramite induzione da substrato (il lattosio).
L'operone triptofano o operone trp è invece un esempio di operone reprimibile, tramite repressione da prodotto finale.
In questo caso infatti, la presenza dell'aminoacido triptofano all'interno della cellula determina il blocco della sintesi degli enzimi che lavorano in modo coordinato per la biosintesi del triptofano stesso.
Il repressore del triptofano si lega all'operatore solamente nel caso in cui sia legato al triptofano. Il triptofano presente agisce da corepressore (repressione da prodotto finale), quindi il grado di espressione dei geni per la sintesi del triptofano è inversamente correlato alla presenza di questo aminoacido nel mezzo di coltura.
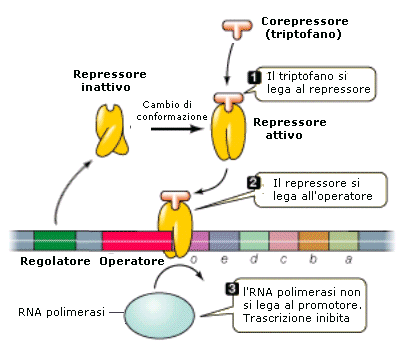
Negli eucarioti il controllo dell'espressione genica è più complesso e le nostre conoscenze a riguardo sono ancora molto incerte. Si ritiene che negli eucarioti l'espressione genica avvenga su più livelli e non solo a livello trascrizionale, come avviene nei procarioti. Si ritiene, ad esempio che le proteine nucleari (istoni e non istoni) possano giocare a questo proposito un ruolo importante.
|
| Appunti su: cellula eucariote animale, cellula eucariota animale, membrana nucleare con anse collegate al reticolo endoplasmatico, giunzione occludente, cellula eucariote, |
|
| Appunti Geografia |  |
| Tesine Ingegneria tecnico |  |
| Lezioni Biologia |  |